 今朝の新聞記事でも南スーダンの内部抗争(Dinka-ディンカ族とNuer-ヌエル族)が報じられていますが、
折角、セム系イスラム教徒が支配する北スーダンからネイティヴ・アフリカンの国として独立したにもかかわらず、
次はネイティヴ・アフリカン同士の対立になってしまった。
何でアフリカ大陸はこうも民度が低いのだろう?ダゲスタン等コーカサス一帯も今だもって地元民族同士の抗争が絶えない。
今朝の新聞記事でも南スーダンの内部抗争(Dinka-ディンカ族とNuer-ヌエル族)が報じられていますが、
折角、セム系イスラム教徒が支配する北スーダンからネイティヴ・アフリカンの国として独立したにもかかわらず、
次はネイティヴ・アフリカン同士の対立になってしまった。
何でアフリカ大陸はこうも民度が低いのだろう?ダゲスタン等コーカサス一帯も今だもって地元民族同士の抗争が絶えない。両者ともかつてのY-DNA「R1b」と「R1a」の西欧列強が支配した地域です。住民を低い民度に抑え込んできたつけが 今頃回ってきているのかもしれません。 それなら南スーダンを更に西と東に分割してしまえばよさそうなものですが、下記のリストにあるように南スーダンには 他にもShilluk族やLuo族等かなりの数の部族が共存しているため 事は簡単ではないのです。部族単位の辺境国家になってしまう可能性が大なのです
かつてイスラム教はサラセン帝国などアラブ民族が大帝国を樹立し当時最先端の文明を持ち、中東から北アフリカの遺伝子分布を すっかり変えてしまったのですが、 栄枯盛衰の理の通りに現在は衰退しています。しかし一時衰退した中国が再度上昇に成功したのは、多種のY-DNA/mtDNAの遺伝子 ハプロタイプ交配による集団エネルギーの向上に成功したからですが、 イスラム圏は残念ながらある特定の少数ハプロタイプ遺伝子しか存在しないため、集団エネルギーが低いままのようです。
この地球の集団エネルギー理論では、生物集団は多様性を増す方向に進化するように設計されており、純系を保つことはそれに 反するのですが、 人類も同じで残念ながらコーカサスも中東もアフリカも民族や文化や遺伝子の純系を維持する方向に向いているようです。
何故、抗争が絶えないのか? 当ガラパゴス史観の記事3-2.日本人のガラパゴス的民族性のrev.5でご紹介した「政治の起源」でも アフリカのことに触れていますが、 いまだに古代的な血縁=言語で集団化した血縁言語部族社会であることが弊害の最大の要因のように思われます。欧米や日中韓のような 極東ではかなり昔に克服された「部族」が、 信仰する宗教も加わり統治の最上位に存在するのがアフリカや中東です。
では部族とは何か?日本列島でも蝦夷族、隼人族、熊襲族などの縄文部族がかつて存在しました。吉備族や出雲族のような 長江文明系弥生部族も中国地方には存在しました。 しかし朝鮮半島から半島内の生き残り抗争に敗れ追い出され、武装侵攻集団として日本列島に渡来した物部などの天孫族や最後に 勝ち残った大和朝廷族、各地に割拠した武士団族が 縄文部族や弥生部族を征服し君臨したことで強固な古代部族社会はなくなり、大和朝廷族の元に統一されたのですが、 国土が狭く海に囲まれていたからこそ出来た奇跡だったと思われます。
ところが、広いアフリカ大地では、統一国家などできるはずもなく、植民地化されて初めて地図上に引かれた国境線で 植民地域ごとに無理やり統合されたのが実態です。 しかし、ひとたび独立すると元の部族社会の複合体に逆戻りしてしまっているのも実態です。
それでもサハラ以北ではイスラム教が接着剤の役割を果たしていますが、部族/民族間対立の方が弊害が大きく、 アラブの春はこのままだと失敗に終わりそうな様子で、 結局、力を持った部族の強権政治に戻らざるを得ないのか複雑な思いです。
サハラ以南では接着剤のイスラム教が弱く、植民地化したヨーロッパ国家が持ち込んだキリスト教も強力な接着剤にはなれず、 強力な古代的な血縁=言語部族集団が復活し強権発動し、 抵抗する弱者部族と虐殺を繰り返す結果になっているようです。
と言うような、現実を踏まえてY-DNA遺伝子調査をまとめて見ましたので、ネイティヴ・アフリカンに興味のある方はご覧ください。 また記事15-12.でご紹介したように、 我々日本人から見ると皆同じ黒人に見えるネイティヴ・アフリカンですが、アフリカ以外の全世界の全ての出アフリカした 非ネイティヴ・アフリカ人を合わせたより、 ネイティヴ・アフリカンの方が遺伝子的に多様なのです。そのことは記事15-12.のNatureの紹介記事に書きましたし、 3-1.で簡略化したY-DNAツリーでもご紹介したので合わせて見てください。
我々の直接の先祖「出アフリカ組」は6万年前頃のホモ・サピエンスの一部の2000人程度が酷暑の大地となったサハラを逃げ出し中東に渡り、 兄貴分の先住ネアンデルタール人と交配をしながら分化し、インド亜大陸で更に細分化し世界に拡がったのです。
ホモ・サピエンスY-DNA「A」とmtDNA「L0」はアウストラロピテクス(猿人)の発祥の地とされる東アフリカからアフリカ全土に 拡散をしたわけですが、サハラの砂漠化後はアフリカ大陸南部のナミビア周辺に居住しています。
その時に人類がまだチンパンジーの祖先と分離し始めた頃に住んでいた森林地帯(中部アフリカ)に居住したY-DNA「A」集団の一部が、 森林地帯での生活に適した小柄なアウストラロピテクス時代の大きさ(ピグミー状態)に逆戻りしY-DNA「B」となり、 またその一部の2000人程度が先輩人類の原人(エレクトス)や旧人(ネアンデルタール人)と同様に出アフリカを決行し、 中東で絶滅寸前のネアンデルタール人と交配し、 ネアンデルタール人が出アフリカ後のヨーロッパ大陸や中東で数十万年掛けて獲得した形質や文化、技術を一気に獲得し、 「出アフリカ組」Y-DNA「DE」と「CF」に分化したのです。
ところがアフリカ大陸では予想しないことが起きたのです。出アフリカした我々の祖先のうち、我らが縄文人のY-DNA「D」の 兄弟ハプロタイプのY-DNA「E」が「出戻りアフリカ」をしてしまったのです。そして「E」はあっという間に全アフリカ大地に展開し、 「A」や「B」と共存し、今や純系の「A」部族や「B」部族は存在しないほどです。 しかし圧倒的に人口の多い「E」遺伝子には純系部族が存在しています。
そして出アフリカ組より遺伝子的に極めて多様なネイティヴ・アフリカンが血縁=言語部族を越えて統合しつつあるのが皮肉にも 奴隷として送り込まれたアメリカやカリブ海諸国や旧植民地の宗主国です。 故国と切り離されたネイティヴ・アフリカン達はアフリカ系移民として移民先の言語を母国語とする新たな集団を形成しています。
民度が低いままの故郷のアフリカでは虐殺をし合うような敵対部族出身者達も、民度の高い新たな故国では ネイティヴ・アフリカンとして新たな1つの社会集団にまとまってゆくのではないかと思えます。 下記の部族リストの一番最後の行がアメリカのアフリカ系国民のY-DNA頻度分布になります。 Niger系のY-DNA「E1b1a」の部族が、中でも特にY-DNA「R1b」の頻度の高い部族が、数多く奴隷として連れてこられたことを物語っています。
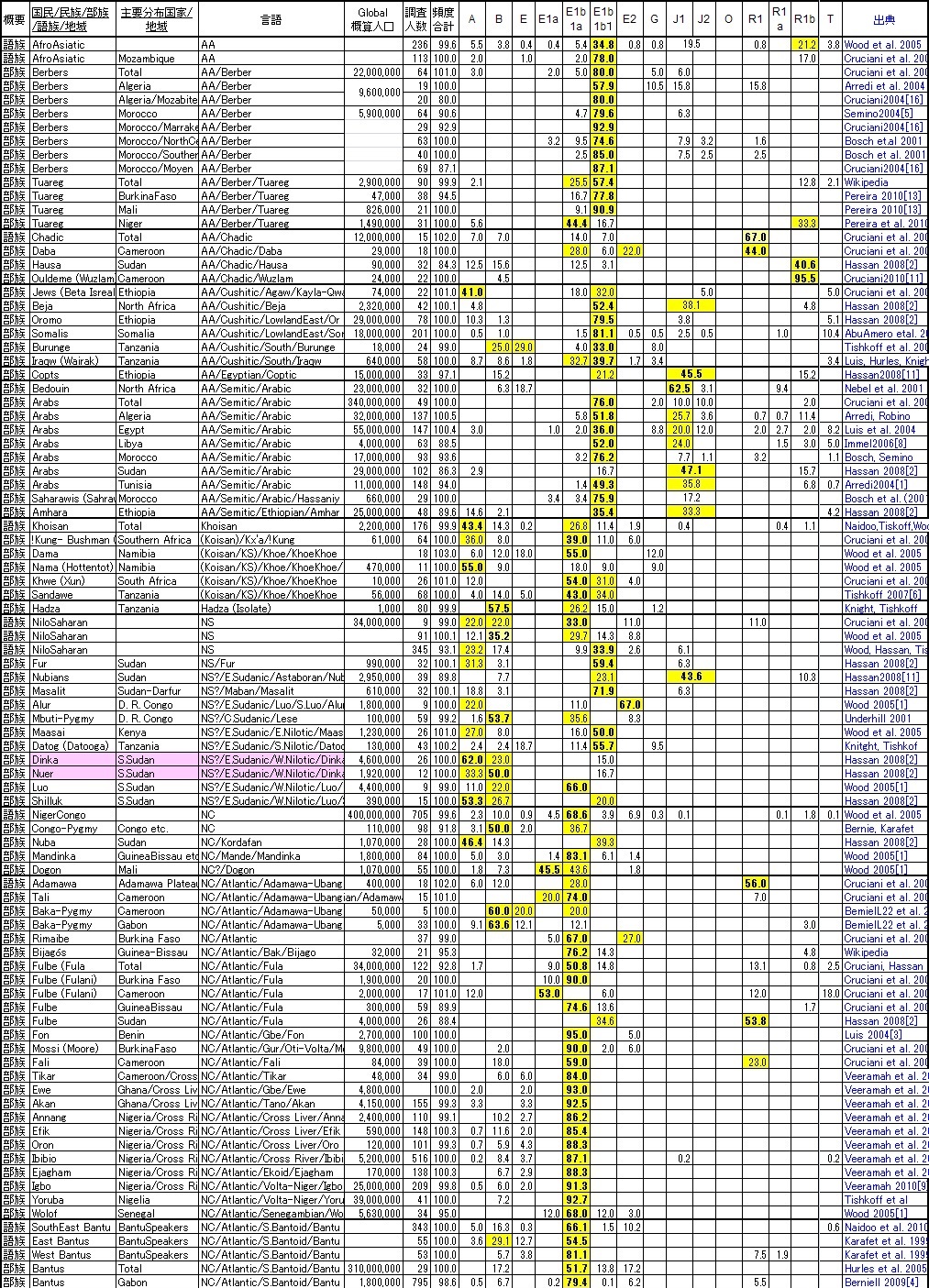
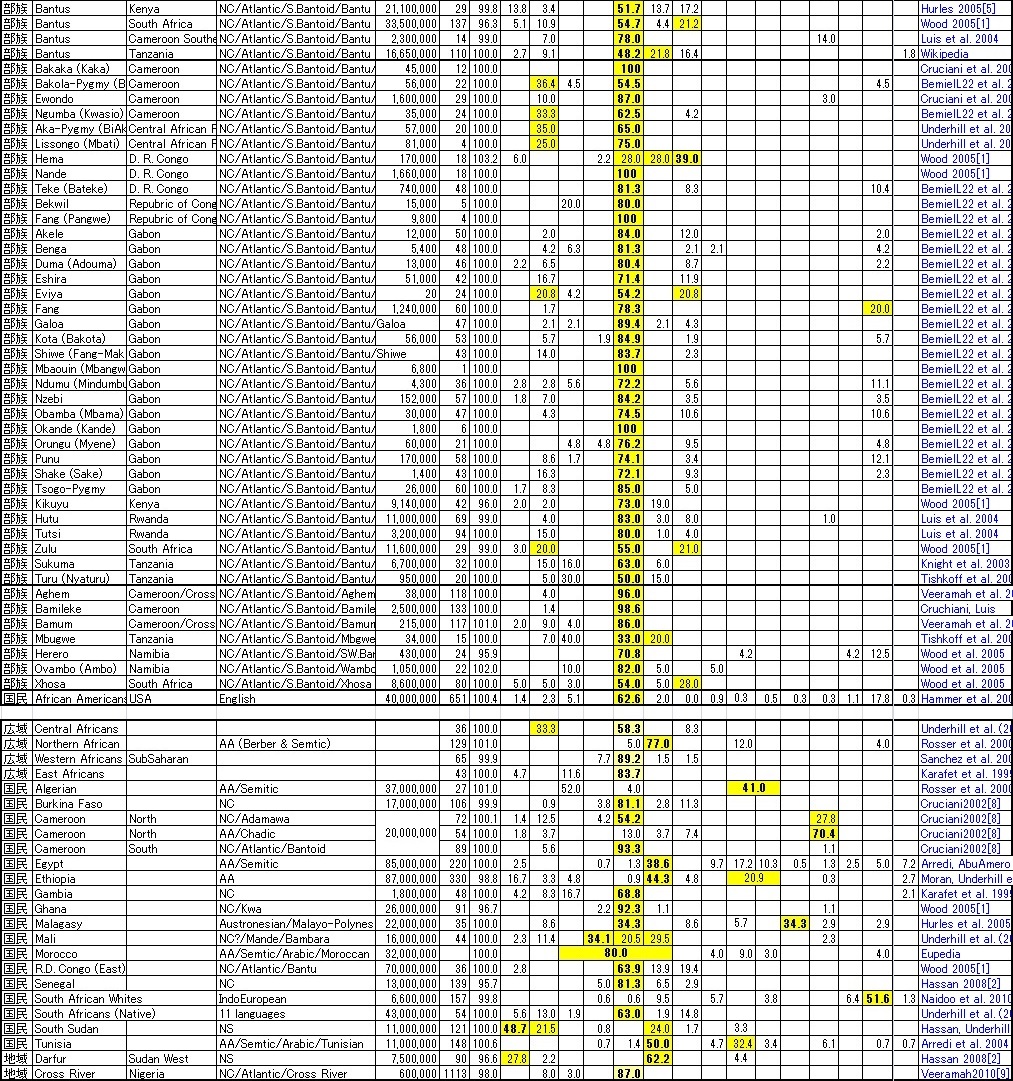
この結果は血縁=言語部族であることを見事に示していると言って過言ではありません。言語族分類はまだ完全に決定している わけではないようですが(そもそも日本語だってウラル・アルタイ語族ではなく孤立言語のようです)、 有力な分類では、現在アフリカの4つの言語群と1つの希少言語は以上のリストの通りです。
1.ホモサピエンス発祥時の遺伝子を維持するのがKhoisan語族:
起源遺伝子Y-DNA「A」を核に、出アフリカしたにもかかわらず出戻ったNiger系遺伝子「E1b1a」が完全共存しています。
最も古い遺伝子集団になります。
2.Dinka族やNuer族が属するNiloSaharan語族:
Y-DNA「A」に加えて、「B」,「E1b1a」,「E1bib」が共存しています。古さを併せ持っています。ただし言語族として成立するかはまだ
疑問符?のようです。将来分類が変わる可能性があります。
3.セム系が属するAfroAsiatic語族:
見事にヨーロッパのラテン系遺伝子のY-DNA「E1b1b」の世界です。ただしセム系になると征服者のY-DNA「J1」が主役になります。
4.最も属する部族が多く、Bantu族も属するNigerCongo語族:
セム系を除くアフリカンとしては最大の言語族で、バンツー族が中心のY-DNA「E1b1a」の世界です。
Y-DNA「A」の次に古い起源遺伝子Pygmy系Y-DNA「B」のピグミー族もNigerCongoに属すらしいです。
5.今のところ孤立言語とされるHadza語:
Khoisan語族に入れる言語学者もあるようですがPygmy系遺伝子Y-DNA「B」が核の部族である以上、
「A」が核のKhoisanに分類するのは遺伝子から見ると無理がありますが、「E1b1a」に征服され変質したと考える余地があります。
日本列島で現アイヌ民族は、Y-DNA「D2」原アイヌがY-DNA「C3c」オホーツク文化の古代ニヴフ族に征服され、
古住シベリア族文化にすっかり変質してしまったのと同じような現象かもしれません。
意外だったのは世界をリードするY-DNA「R1b」がネイティヴ・アフリカンにも入り込んでいることです。 特にAfroAsiaticのChadic系は「R1b」の要素が大きく入り込んでいて、ヨーロッパではケルトやバスクになった遺伝子集団がアフリカでは Chadic集団になったようです。
こうなると、他の記事でも書いていますが、見かけで分類された「人種」という概念は改めて無意味ではないかと思えます。 人間がいかに「外観」で判断をしているか、と反省があります。
我々Y-DNA「D2」縄文系の子孫に取って世界で最も近縁な集団は同じY-DNA「D」ハプロタイプで、羌族とチベット族の中の Y-DNA「D1」集団と 最もオリジンなアンダマン諸島のY-DNA「D*」のJarawa(ジャラワ)族とOnge(オンゲ)族ですが、その次に近縁なのは ネイティヴ・アフリカンやラテン系のY-DNA「E」集団なのです。かつてはY-DNA「DE」のことを「YAP」と呼んでいました。
Y-DNA「DE」に共通の特徴はY-DNA「CF」系と異なり、ネアンデルタール人の形質をあまり受け継いでいない、ということです。 「CF」系は大柄・色白・紅髪・彫深顔..............出アフリカしたネアンデルタール人が中東・ヨーロッパの 高緯度地域で数十万年掛けて獲得したエピジェネティクス(後天的獲得形質)を交配によって文化・技術等と共に一気に獲得したのです。
中国の黄河周辺古代遺跡から発掘される人骨は明らかにコーカソイドの特徴を持っていることは、西欧の人類学者の中では定説のようです、 つまりY-DNA「CF」の子亜型であるY-DNA「O3」は本来彫深顔だったのですが、黄河流域の「黄砂」にさらされる過酷な環境で黄砂適応という エピジェネティクスを獲得しフラット顔に「進化」したというのです。
極東ではこの他に寒冷化したシベリア大地に住む古住シベリア系住民が獲得した寒冷地適応のエピジェネティクスもあります。 Y-DNA「C3c」,Y-DNA「N」やY-DNA「P」等が寒冷地適応のフラット顔を「獲得」したようです。
また、Y-DNA「J2」のメソポタミア農民系と共にメソポタミア農耕文明を築いたとされるアルプスのアイスマンやスターリンでお馴染みの コーカサス系のY-DNA「G」が、少数の出現頻度ですがアフリカ大陸に農業を持ち込んだことが見てとれます。
また、第3代アメリカ大統領のジェファーソンの遺伝子Y-DNA「T」もアフリカ大地で少数ですが広い地域で出現します。 Y-DNA「A」,「B」,「E」が主役のアフリカ大陸ですが、脇役もしっかりと存在しています。
マダガスカル島(マラガシー)だけに存在するのが、長江文明系Y-DNA「O1a」(楚系?)と「O2a」(越系?)です。 いつ頃どうやって渡ったのかはまだ未調査ですが、日本人にも存在するY-DNAです。こんなところに同族遺伝子がいるとは驚きです。 しかも両者合わせてマダガスカルの34%を占めているのです。
中東では絶対の出現頻度トップを誇るセム系Y-DNA「J1」が、北アフリカのアラブ社会では地中海・ラテン系のY-DNA「E1b1b」より 頻度が低いことも意外でした。北アフリカのイスラム・アラブ社会は一見中東のそれと同質に見えるのですが、実は現地の先住民である Y-DNA「E1b1b」を征服した結果であったことが良くわかります。
特にチュニジアはローマ帝国時代からハンニバルの活躍等で知られる典型的なY-DNA「E1b1b」の地中海・ラテン国家だったのですが、 Y-DNA「J1」のイスラム・アラブ人の進出で完全に変質してしまい、今やイスラム・アラブの一員になりきってしまったのです。 それでもY-DNAを調査することで本来北アフリカはY-DNA「E1b1b」世界であったことが理解できるのです。
この調査のきっかけとなった南スーダンのDinka族とNuer族は言語はほとんど同一になり今では方言程度の違いしかないのですが、 遺伝子調査の結果は、Dinka族がY-DNA「A」を核にするコイサン起源、Nuer族はY-DNA「B」を核にするピグミー起源であることがわかりました。 もともとの遺伝子レベルでの起源の違いのため違う言語=部族社会であったものが、ラテン系Y-DNA「E1b1b」に統治され、 その後セム系の征服者によって長い間統治された間に交配が進み一見同質になったかのように見えたものが、 セムの「重し」が取れた途端に本来の異なるオリジンが顔を見せたようです。
以上、アフリカ大陸は起源遺伝子Y-DNA「A」と「B」を古代遺伝子Y-DNA「E」が征服し古代部族王国を形成してきたところに、 西欧列強が植民地化し、古代血縁=言語部族分布を無視して地図上で勝手に国境線を引き部族を分断し、 そのまま独立したため今でも部族が複数国家間に分かれて居住しています。
日本のように縄文系も弥生系も武装侵攻系も今では既に意識がないくらい交配が進み「日本民族 (アイヌ系の統合はもう少し時間がかかりそうですが)」がほぼ確立されると部族対立はもうありませんが、 韓国では征服者の中華王朝出先機関の子孫系が多い中部・北部と長江文明系農民の子孫が多い南部の部族社会、 扶余系のツングースの子孫が多いはずの北部の名残が、大統領選挙等で話題になる「××道」の地域特性にまだ少し残っていますが、 そのうちなくなり一体化するでしょう。
北朝鮮は全くデータがないため見当がつきませんが、中国の国境周辺の朝鮮族のY-DNA「O2b」長江文明系の頻度は韓国(30%)より高く(43%)、 中華王朝出先機関系Y-DNA「O3」は韓国(43%)より低い(32%)のですが、ツングース系Y-DNA「C3c」はほぼ同じ(11~13%)で基本構成は同じなので、 地理的に韓国と中国朝鮮族の中間と考えてよいでしょう。 このように朝鮮族は3つの国家に分断されている事実は厳然と存在しているのです。アフリカで起きていることが隣の朝鮮半島でも 起きているのです。似ているようで似ていない日本人との最大の違いはY-DNA「D2」,「C1a」,「C3a」の3縄文系亜型が朝鮮・韓国では完全に 抜け落ちているところです。
最後に、Y-DNA「E」系ネイティヴ・アフリカンは日本人のY-DNAの40%以上を占める縄文系Y-DNA「D」はY-DNA「DE」から分化した、 兄弟遺伝子なのです。
以上
表紙に戻る