最新の研究では、最初のホモサピエンスであるミトコンドリアイヴ(mtDNAイヴ)の出現は20万年前頃らしいです。 当然のことですが原人の中に一人のホモサピエンスが誕生しても今の現代人類が生まれたわけではありません。 そのmtDNAイヴが進化していない原人男性と交配しながら進化した遺伝子を持つ子供を産み、その子供たちが進化していない相手と交配し、 少しづつホモサピエンスの遺伝子を広げ、13万年前頃にやっと男性のホモサピエンスのY-DNAアダムが誕生したいうことなのです。つまり進化は女性から始まり、男性に伝播してゆくのです。
これは当たり前で本来生物は「メス」のみの単性生殖でしたが、それではバクテリアからバクテリアが生まれるだけで生物は永遠にバクテリアだけだったのですが、 生殖専用の「オス」という存在が「メス」から創り出され、生物を多様化させることに成功したのです。この「オス」の創造が地球にとっては最大の発明なのでしょう。 と言う訳で、数十億年かかって人類まで到達した、という事だそうです。
mtDNAイヴが誕生してY-DNAアダムが出現するまで7万年近くかかっているというのですが、これは人類にとっては長い期間ですが、 生物の歴史の中では一瞬でしかありません。同時出現と同じなのです。
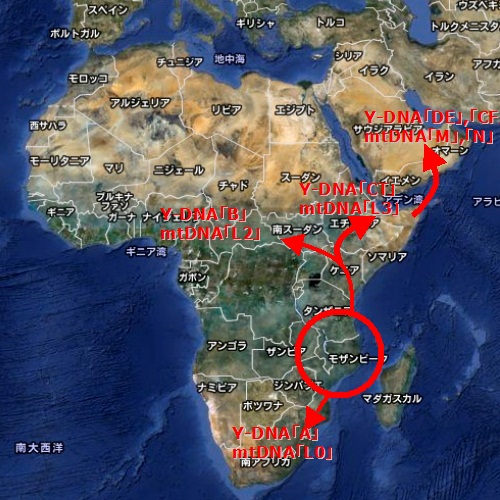
こうしてY-DNA「A」が誕生しmtDNA「L」と共にアフリカ大陸の今でいえばケニア、タンザニア辺りの森林地帯で少しづつ増殖をしたようです。 この頃のY-DNA「A」は当然ですが草原でアウストラロピテクス時代と変わらない狩猟をおこなっていたようです。
ところがアフリカ大陸は徐々に灼熱化し、スタンフォード大学の研究では、2000人ほどの絶滅危惧種に陥り、先輩人類と同様新天地を求め出アフリカを決行したらしいのです。 研究者は出アフリカしたのは今の角アフリカと言われるソマリア近辺だろうと考えています。当然ですが、海面は今より低くアフリカ大地とアラビア半島はこの辺りで繋がっていたのです。 そしてこの中東で人類がであった最大の出来事は亜種で先輩人類のネアンデルタール人との出会いと交配でした。 60万年ぐらい前に先に出アフリカしたネアンデルタール人は既に当時の新人と比べるとはるかに高度な文化や技術を持っており、更に北欧付近で高緯度地適応もしていた様です。 亜種間交配した新人は、ネアンデルタール人が数10万年かかって獲得したすべてを、一気に獲得したのです。
Stanford大学の研究で、我々現代人の祖先は60000年-70000年前頃の気候大変動で2000人程度まで減少し絶滅する寸前の「絶滅危惧種」だったそうなのですが、 そのことが原因で生き延びるために現代人の祖先は必至の思いで、全く住めなくなった極砂漠化したアフリカ脱出を決行し、 当時陸続きだったアラビア半島南端からサバンナ気候で住みやすかった中東に逃げました。 そこには先住民族のネアンデルタール人がいて多少の交配があったようです。その頃の気候図がWikipediaにありました。
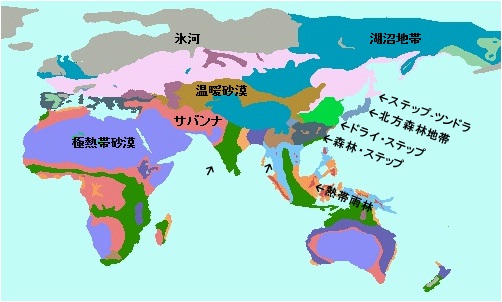
我々出アフリカした現代人の遺伝子の1-4%はネアンデルタール人との交配の結果取り込まれたものだそうです。 そのサバンナ気候の中東にどのくらいの期間住み着いたかは詳しくはわかっていませんが、オーストラリア亜大陸に アボリジニの祖先が現れたのが50000万年以上前と明らかになってきたことは、現代人類の移動は早かった、ということです。 80000年前には中国大陸に到達していたという最新報告もあるくらいです。 当時の海岸線沿いにひたすら移動を続け陸続きの最西南端に一気に到達したようです。このためアボリジニは古い形質を残していると考えられています。
このように最終氷期当時の海岸線は現在よりも最大で120mも低かったと考えられています。 このため現在ユーラシア大陸の沿岸にある島はアンダマン諸島もインドネシアの各諸島も海南島も台湾も日本列島もサハリンもすべて陸続きのだったようです。
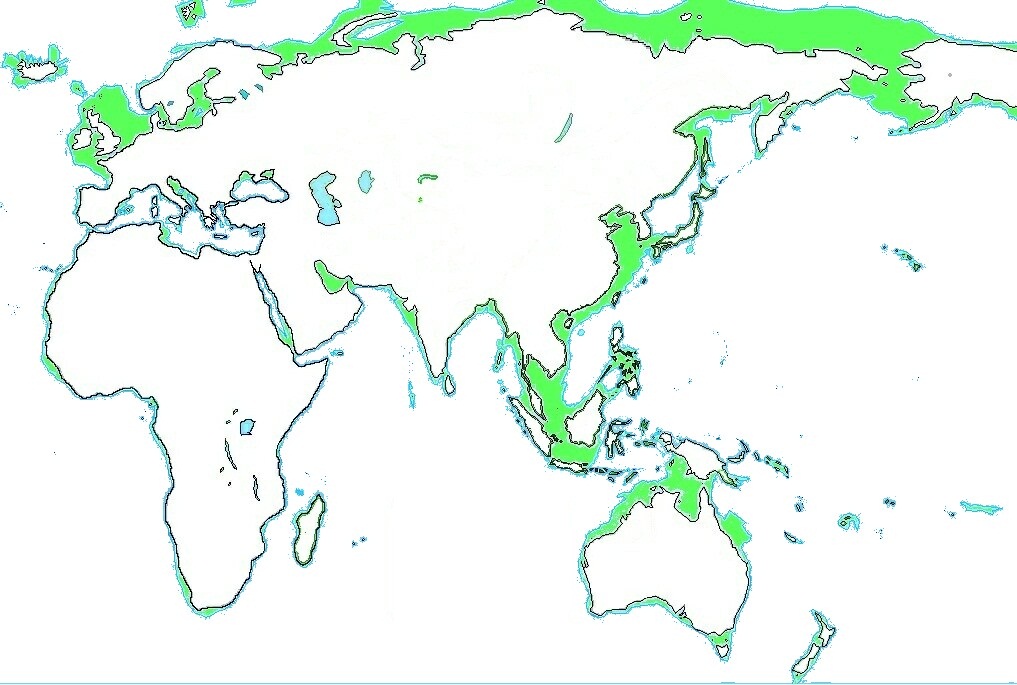
2つの地図はすべてWikipediaから拝借したものですが、上の地図は最終氷期の最寒冷期の陸地をあらわしているそうです。 一方下の地図は上図に現代の陸地を黒い線で示し、、当時ユーラシア大陸の沿岸部で現在海没し大陸棚になった部分を緑色で示しています。 これで一目了然なのは日本列島もユーラシア大陸の一部でアメリカ大陸まで陸続きだったことです。 こうしてアボリジニの祖先は出アフリカ後一気に陸地の最東南端であるオーストラリア亜大陸に到達したそうです。
では我々日本人の最大の祖先である縄文人の祖先集団は出アフリカ後Y-DNAハプロタイプ「D」が確立した55000年前頃から縄文文化草創期の11000年前頃まで、 どこで何をしていたのでしょうか?まだ全くわかっていません。チベットにY-DNA「D1」「D3」が存在するということは 日本列島弧部分に到達する前に亜型に分化する時間があったと考えるのが自然です。 同様に「C」もニュギニアに亜型の「C2」、アボリジニの「C4」があることからオーストラリアに到着する前に既に亜型に分化していたと推測できます。 このためスンダランドがその分化の場所と今は考えられているようです。
以上
