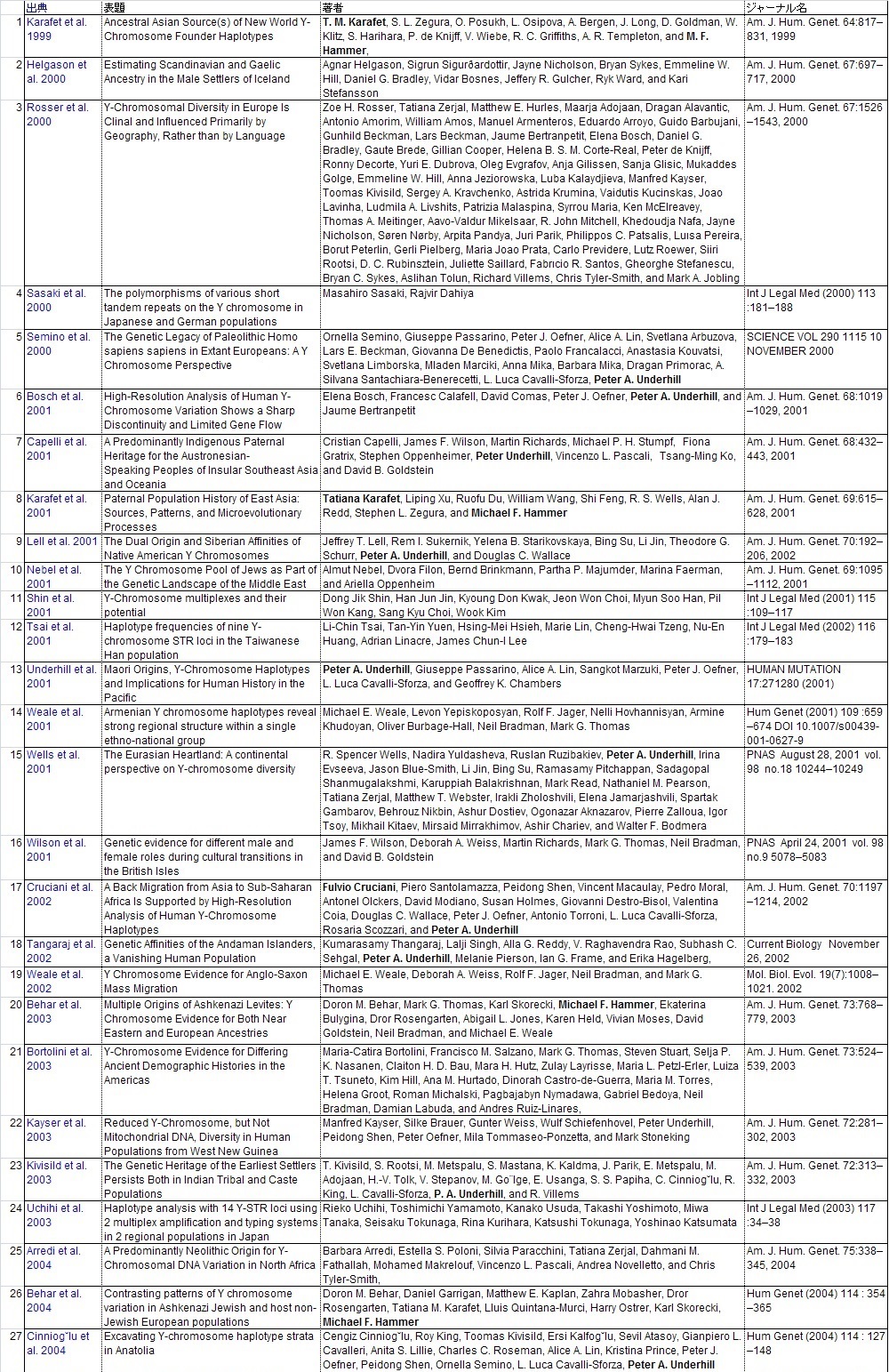
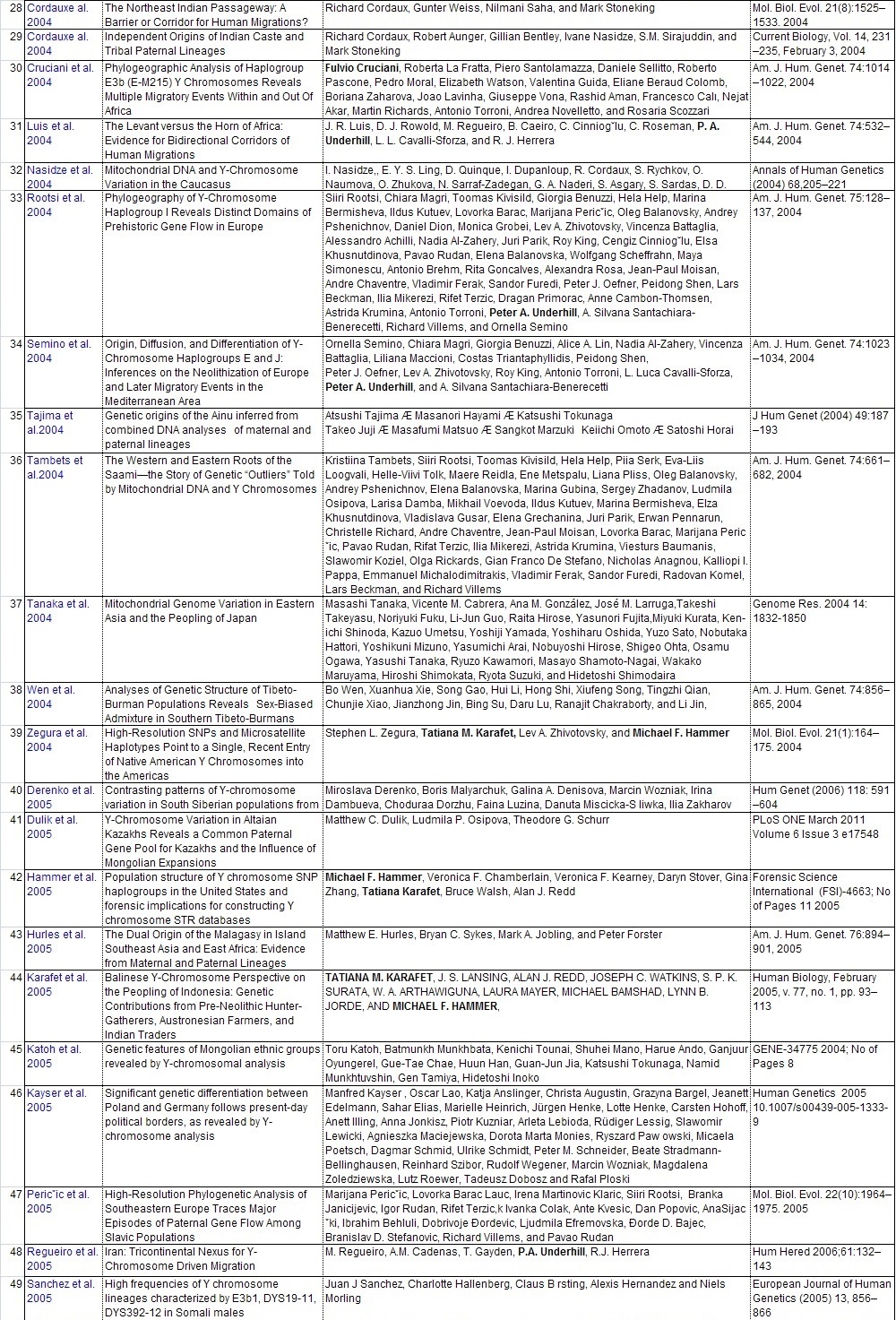
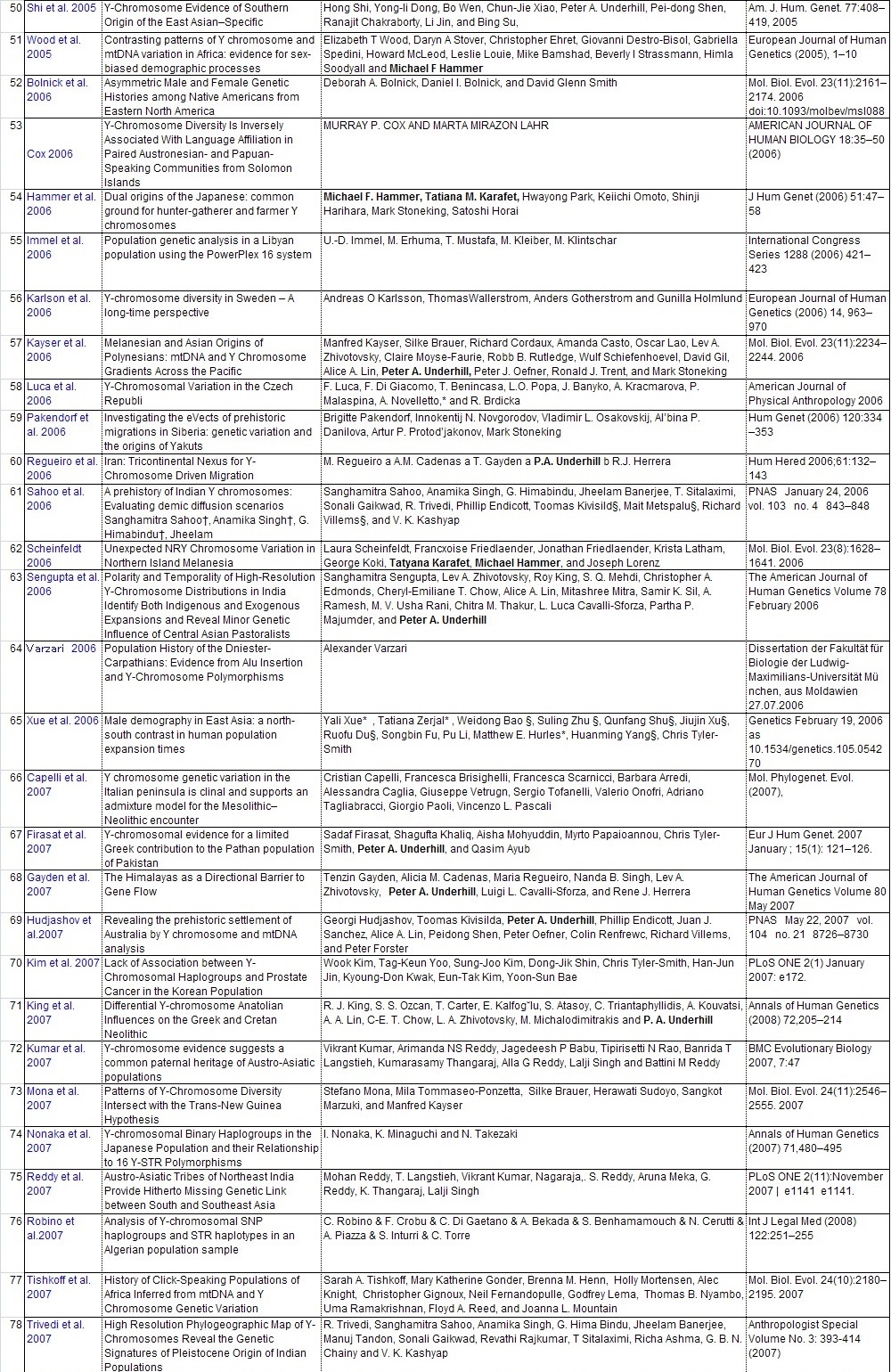
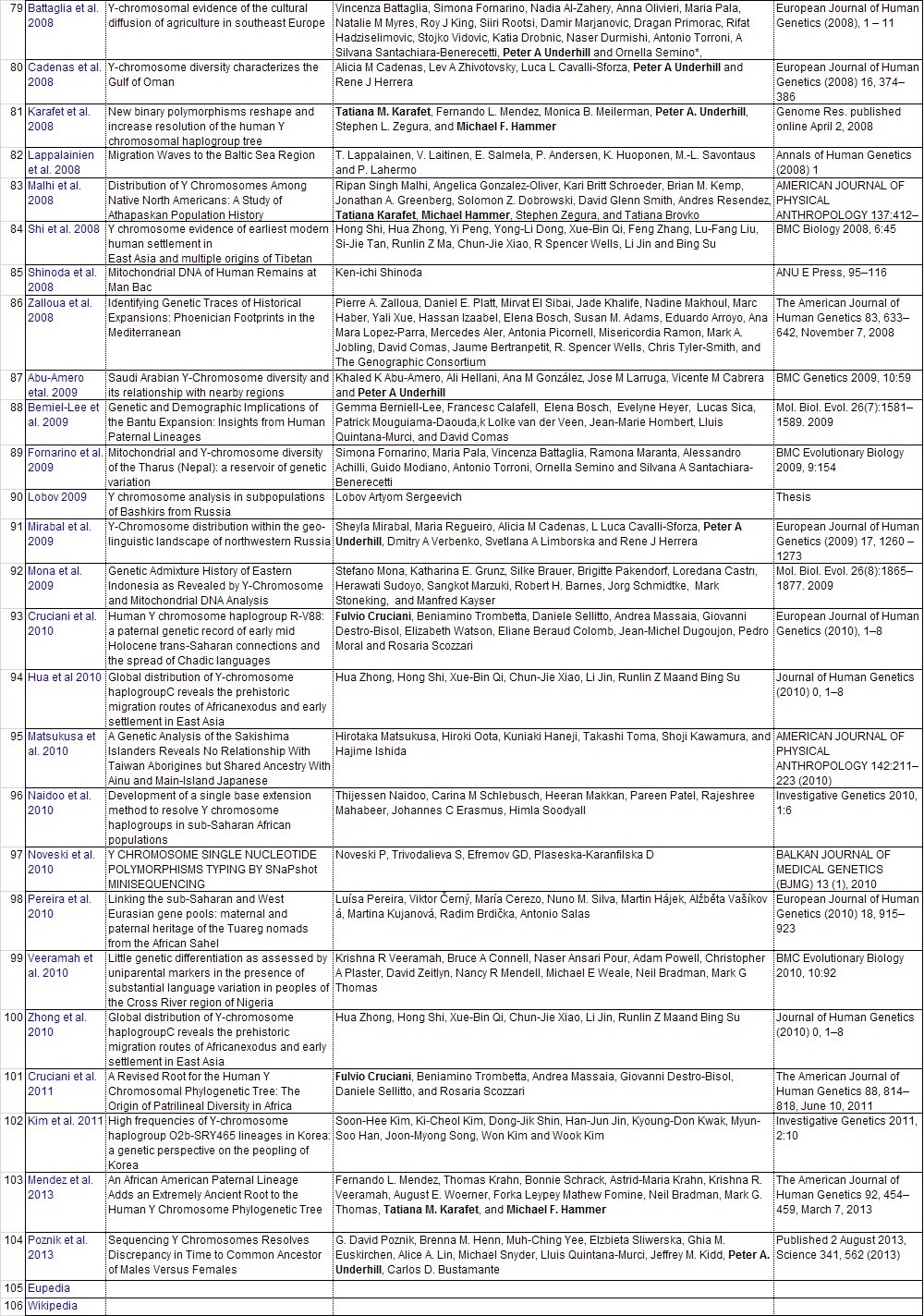
Y-DNAのハプロタイプの頻度集計と解釈に直接役立った104論文のリストを添付します。ご自分で調査し ご自分の史観を確立したい方は、是非読んでください。 また各論文の参照論文を更に調査することで当ガラパゴス史観以上のデータが集まる可能性があります。
他人の説は当てにせず、自分で調査をすることが史観形成の第一歩です。枝葉末節はさておき幹を作りましょう。 批判する人は枝葉末節を問題にしますが、大所高所から俯瞰的に見る姿勢が必要です。 多量のデータからデータマイニングしそこから史観を導き出しましょう。 データの結果の解釈は人それぞれです。是非あなた独自の史観を形成してください。
以上
表紙に戻る