Y-DNA及びmtDNAの論文104編を読み込みデータマイニングし統計解析した結果得た推論に基づき、 ガラパゴス史観を総括した日本人の源流考です。 追加の着想がまとまる都度書き足します。
V.1.92 mtDNAの新しい解釈を付け加えました。ご参考に。
V.1.91 この項の最下段に、Y-DNAの日本人の源流ツリ−をアップしましたので、ご参考に。
v.1.9 そろそろ終活に入りましたので、全編を簡潔に書き直し中です。間違いなども併せて修正中です。
内容
1. はじめに、素朴な疑問から
2. 草創期のネアンデルタール人(ハイデルベルグ人)から始まったようだ。
3. 原ホモサピエンスから現ホモサピエンスへ脱皮したのではないか!
3-1. 華奢型への進化(?)と集団形成への変化があったのではないか!
3-2. 最新のY-DNAツリーの超概要!
3-3. 現代人類はホモサピエンスと終末期の成熟型ネアンデルタール人とのハイブリッド?
3-4. 現代人類は進化の爆発に遭遇できたのではないか!!!
4. 日本列島への最初の到来者は、古代遺伝子集団:Y-DNA「D」と「C」
4-1. 縄文系遺伝子集団:Y-DNA「D」の移動
4-2.縄文系ハンター遺伝子集団:Y-DNA「C」の移動
4-3. Y-DNA「D」の先輩チベット人の高高地適応獲得形質:
4-4. Y-DNA「D」の弟亜型のY-DNA「E」とは?
4-5.縄文人のコアをなすY-DNA「D1b」,「C1a」の日本列島への上陸
4-6. さらにY-DNA「C2a」の日本列島への上陸
5. 長江文明系稲作農耕文化民の日本列島到来
5-1. 長江文明系稲作農耕文化民の拡散
5-2. 日本列島と朝鮮半島の稲作農耕民の共通点
6. 黄河文明系武装侵攻集団の到来
7. 日本人の源流....簡易まとめ
8. Y-DNAの分化.....エピジェネティクスか"
9. 雑考
10. 時代の趨勢
11. 余談
1.はじめに、素朴な疑問から
Y-DNAとmtDNAの分化ツリーの調査を進めて行く過程で下記のような疑問が生じました。
(1)アフリカ大陸に居住するホモサピエンス始祖亜型のY-DNA「A」と「B」は、なぜ狩猟採集の形態から変化しなか
ったのか?
(2)始祖亜型Y-DNA「B」が出アフリカ後、分化した古代4亜型の中でY-DNA「D」,「E」,「C」の、アンダマン諸島
、サハラ以南のアフリカ大陸、オーストラリア、ニューギニアなどの僻地に残った古代亜型も「A」,「B」同様
、なぜ狩猟採集の形態から変化しなかったのか?
(3)変化したのは古代4亜型の中でインド亜大陸に落ち着いたY-DNA「F」だけだったのはなぜなのか?
(4)狩猟採集形態を維持し続けた各地の古代亜型遺伝子集団は、解剖学的現代人類(現ホモサピエンス)として完成
していたのだろうか!?
(5)始祖亜型Y-DNA「A」,「B」は原ホモサピエンス、古代亜型Y-DNA「C」,「D」,「E」は実は“古ホモサピエンス”
の段階にとどまっていたのではないか?
(6)古代亜型Y-DNA「F」から分化した、Y-DNA「G」から「T」までのすべての現代亜型の分化地域はインド亜大陸
ー中央アジア周辺に集まっている。最後に分化したY-DNA「R」も、最も有望な起源地は中央アジアあるいはロシ
ア/シベリアの南部辺りと考えられている。このことは古代亜型Y-DNA「F」の子亜型の現代亜型群が完成形“現
ホモサピエンス”に変化した、と解釈すると合理的ではないのか!?
(7)ホモサピエンスはアフリカ大陸で完成形になり出アフリカしたと思い込んでいるが、始祖亜型Y-DNA「B」の出ア
フリカ後にネアンデルタール人と遭遇しハイブリッド化することで解剖学的現代人類が完成したと考えられない
か?
(8)古代亜型Y-DNA「F」のみが現代亜型群に分化したのは、インド亜大陸と周辺地域のネアンデルタール人(デニソ
ワ人も含む)とのハイブリッドによる、と言っても良いのではないか?!
2.草創期のネアンデルタール人(旧名ハイデルベルグ人)から始まったようだ。
ホモサピエンスの亜種か異種かとこれまで議論されてきたホモ種のネアンデルタール人は、最新の欧米の遺伝子解析の知見では、 80万年前頃にホモエレクトスから草創期のネアンデルタール人(旧名ハイデルベルグ人)が出現し、ネアンデルタール人とホモサピエンスの 共通の祖先と考えられるようになってきています。ネアンデルタール人は60万年ぐらい前には出アフリカし、先輩人類としてユーラシア大陸に 拡がり地方型に分化し、そして40万年前頃にネアンデルタール人のシベリア型のデニソワ人が地方型として出現したようです。
注:ホモエレクトスとネアンデルタール人との間の古人類種と分類されてきたハイデルベルグ人は、最新分析技術によ
る遺伝子解析の結果、独立古人類種と考えるより草創期/初期もしくはプレネアンデルタール人と考えるのが妥当
だと、分類提案されることになりました。このため60万年前頃に出現したと考えられてきたネアンデルタール人
の出現年代は、ハイデルベルグ人の出現年代とされていた80万年前頃に遡りました。
注:デニソワ人は遺伝子解析の結果、あくまでネアンデルタール人に包含される地方型というのが極最新の見解です。
ユーラシア大陸に拡大・拡散したネアンデルタール人は数十万年の歴史の中で亜種に近いくらいの各地域型に進化
を遂げていた、と欧米の研究者は説明していますが、あくまでネアンデルタール人の範疇に入るのだそうです。
ともあれ最新の遺伝子解析技術は、現代人類の遺伝子の2−4%はネアンデルタール人から受け継いでいることを解明しました。 東アジア人の比率が最も高く、次にヨーロッパ人で、東南アジア人は意外に低いのだそうです。
特にNatureの論文集「15-28. 古代に移動したDNA−縄文人の先祖の移動も見えてくる!?」では、 Y-DNA「C1b」のパプアニーギニア人とオーストラリアアボリジニは常染色体の研究でデニソワ人遺伝子が5%も受け継がれているのに対し、 海面上昇でサフール大陸がニューギニアとオーストラリアに分裂後にスンダ列島に移動してきた集団(Y-DNA「S」や「M」等)には デニソワ人遺伝子が見つかっていないこと、Y-DNA「D」を持つアンダマン諸島の先住民の常染色体からもデニソワ人遺伝子は 見つかっていないことを報告しています。また解剖学的現代人類が出アフリカしたのは75,000年前より以前、とも報告しています。
また逆にネアンデルタール人にはホモサピエンスの遺伝子が20%も含まれていた、と言う報告まであります。
つまり現代人類は、既にある程度の高度な文化を築き上げていたネアンデルタール人とのハイブリッドの結果、 進化の爆発が起こり、解剖学的現代人類が完成したのではないかと考えるのが妥当なのではないかと思われます。
また、クレタ島の初期旧石器時代の遺物の発見は、ネアンデルタール人がボートを使い、船乗りを習得したことを示唆している 可能性があると報告されています。このことは意外に示唆に富んだ報告です。Y-DNA「C」のニューギニア人やオーストラリアアンアボリジニが 何故スンダ列島を渡りサフール大陸に渡ることができたのか?東海大学の海洋調査でオーストラリアの遺跡からマグロの骨が発掘され、 Y-DNA「C」は海洋性ハンターであることが示唆されていますが、渡海技術は地中海-中近東型ネアンデルタール人から受け継いだ 技術であることがうかがえるからです。つまり、古代亜型Y-DNA「C」はネアンデルタール人とハイブリッドであると考えられるのです。
ホモサピエンスは2段階のハイブリッドである可能性があります。出アフリカした、始祖亜型Y-DNA「B」は中近東あたりで ネアンデルタール人と最初のハイブリッドを形成し、YAP系のY-DNA「DE」と非YAP系のY-DNA「CF」に分化したと考えられます。 YAP系と非YAP系の人類上の源流的な違いに関する調査はまだできていません。なんの意味があるのだろうかはまだ「?」です。 そして非YAP系の古亜型Y-DNA「F」がインド亜大陸周辺で2回目の決定的なハイブリッドを経験し、現代亜型に分化したと考えると最も合理的です。
また大きなトピックスとして、解剖学的現代人類のY-DNA分化ツリーにネアンデルタール人とデニソワ人の亜型がとうとう組み込まれました。 一方、現在の調査結果ではネアンデルタール人とデニソワ人のmtDNA亜型は解剖学的現代人類には今のところ検出されておらず、 ホモサピエンスにはネアンデルタール人のmtDNAは受け継がれていないと考えられています。 つまり、ネアンデルタール人、デニソワ人と解剖学的現代人類はY-DNA(男系)で直系に繋がっている(つまり直系の先祖)、 と言うことになりますが、 女系を表すmtDNAでは断絶している、と言うことになります。
記事「14-2.男系のmtDNAが遺伝しないのは、"ゴミ"だからだった」で解明されたようにmtDNAは母親からしか受け継がれません。 ミトコンドリアはもともと共生したエネルギー産生細菌なので、人類自身のDNAではないのです。恐らくこのために生殖時の染色体減数分裂と 母親、父親の染色体の合体のプロセスに組み込まれないのでしょう。結果、母親側のmtDNAのみが受け継がれると言うことのようです。 (ただし極最新の研究報告では極稀な場合として、男系のmtDNAも遺伝するそうです。)
以上のように、ネアンデルタール人とホモサピエンスは異種か亜種かという問題に対し、ネアンデルタール人のY-DNAの変異型が 特定されホモサピエンスのY-DNAツリーと直系でつながったということは、ネアンデルタール男性人は異種ではないという結論になります。 結果としてネアンデルタール人とデニソワ人の男性は原ホモサピエンスの女性と交雑しても子孫を残せた、あえて言えば、 ホモサピエンスとネアンデルタール人の男性は亜種ほどの違いもなく、恐らく同種で「頑丈型」と「華奢型」の違い程度に過ぎない ということになるかもしれません。
一方女性に関しては異なり、ホモサピエンスの男性とネアンデルタール人やデニソワ人の女性との組み合わせでは 子孫が残らなかったのだろうと、欧米の研究者は考えているようです。残らなかった原因はまだ研究報告がありません。 うがった見方をすればネアンデルタール人とデニソワ人の女性達は華奢でひ弱そうなホモサピエンスの男性を受け入れなかった、 ということも考えられます。
などと考えていたのですが、もっとシンプルに考えると、mtDNAはネアンデルタール人女性もホモサピエンス女性も実は同じだった、 女性は何の変化もしていなかった。とも考えられるのです。もともと「オス」は遺伝子の多様性をもたらすために「メス」を土台に 作られた存在と考えると、土台の「メス」つまり女性は遺伝子変化が起こりにくく、作られた「オス」つまり男性は変化しやすいと 考えると、早期のネアンデルタール人男性から草創期のホモサピエンス男性が遺伝子変異の結果、華奢型体質として突然変異を起こし、 虚弱ゆえに集団を作り弱肉強食のアフリカ大地を生き抜いた。またその集団を維持するために協調性を保てるメンバーのみで集団を構築し、 ボスになりたがるオスや協調性がないオスを排除し集団を強固にし、結果的に同調圧力が強いホモサピエンス体質が形成されたのではないかと 推測で切るのですす。
3.原ホモサピエンスから現ホモサピエンス=解剖学的現代人類へ脱皮したのではないか!
現代人類の祖先は、共通の祖先である草創期のネアンデルタール人からネアンデルタール人が先に出アフリカした後も、 アフリカ大陸に残存していた早期のネアンデルタール人の中から、極最新のアフリカ大陸の調査では、草創期の原ホモサピエンスも 50万年前頃には既に出現していて、30万年前頃にホモサピエンス最古のコイサン集団の祖先が出現し、20数万年前頃には 既に出アフリカが始まり何度か出アフリカを行っており遺跡も発掘され、最近の発掘調査では、10万年前ごろにはすでにレバント地域に 定住していたらしく、8万年前頃には中国南部に到達していたとも報告されています。
しかし6-7万年前頃の最後の出アフリカが、解剖学的現代人類を形成したという解釈になっています。 以前の5−6万年前頃に最終出アフリカしたのではないかという見解が、今は6-7万年頃と遡ってきており、 更に75,000年以上前ではないかという前述の論文まで報告されてきています。
スタンフォード大学の研究では、現生人類の先祖が2000人程度の規模で出アフリカしユーラシアに拡がるまでに、 ネアンデルタール人の歴史は既に60万年以上は経過していたそうです。そしてネアンデルタール人は、3-10万年前頃に 出アフリカ組のホモサピエンスと交雑し、3万年前ぐらいには絶滅した、という見解になっています (デニソワ人の絶滅時期はまだ詳細には論じられていません)。つまり終末期で文化的に最も成熟したネアンデルタール人と ハイブリッドしたということなのです。
3-1.華奢型への進化(?)と集団形成への変化があったのではないか!
いずれにせよ、80万年前に現れた草創期のネアンデルタール人(旧ホモハイデルベルゲンシス)が共通の祖先となり、 60万年前頃にネアンデルタール人が先に出アフリカし、50万年前頃に草創期ホモサピエンスが現れたのだろうと考えられます。 この草創期ホモサピエンスは、恐らく華奢型に突然変異してしまった早期ネアンデルタール人と推測されます。 後述しますが、恐らくこの華奢型に進化?退化?してしまったことが、アフリカ大陸を生き抜くために、 捕食獣から身を守るために集団化せざるをえなくなり、仕事の分担をするようになり、コミュニケーションをよくするため 言語能力を進化させたと、京都大学の松沢教授は説明しています。捕食者の多いアフリカ大陸を生き抜くには、 頑丈な体のほうがダーウィンの自然選択の法則で選ばれるはずです。にもかかわらず華奢なホモサピエンスが生き残ったのは、 集団化することで弱点を補ったのではないかと想像できます。
このことは要するに原ホモサピエンスとは、集団を作ることが本来の生活様式(つまりムラ形成!)ということになります。 最新の論文「14-12. 初期の人間は自分自身を家畜化した、新しい遺伝的証拠 v1.1」報告されているように、人類は同調性のない個体や、 ボスになりたがる個体などグループ維持に不都合な個体を排除し、できるだけ同調できる個体達でグループ構成をするようになった、 と言うことのようです。
日本人の持つ「同調圧力」は、なんと原ホモサピエンス本来の形態ということになります。恐らく家族単位の生活様式だった と考えられるネアンデルタール人には同調圧力も存在せず、数十万年経過しても人口増加率は低かったと思われますが、 集団を形成するホモサピエンスは集団を形成した結果として、人口増加率がはるかに高くなったのではないかと考えられます。
3-2.最新のY-DNAツリーの超概要!
最新のY-DNAツリーは、
Y-DNA「Adam」から
Y-DNA「A0000」(デニソワ型ネアンデルタール人)が分化し、
Y-DNA「A000」(終末期の成熟型ネアンデルタール人)が分化し、
Y-DNA「A00」(最古のコイサン集団)が分化し、更に「A0」、「A」と分化が進みました。
この「A」から「A1」へ、さらに「A1b」へ分化し、
Y-DNA「A1b」からY-DNA「BT」が分化し、
Y-DNA「BT」がY-DNA「B」とY-DNA「CT」に分化しましたが、
このコイサン型「A」とピグミー型「B」はホモサピエンスの始祖亜型で原ホモサピエンスと考えられます。
Y-DNA「CT」が出アフリカし、現ホモサピエンスY-DNA「DE」とY-DNA「CF」に分化しました。
これは中近東あたりで地中海-中近東型ネアンデルタール人との一次ハイブリッドの結果と推測できます。
この中で「C」,「D」,「E」は古代性を強く残した狩猟採集民として、近年まで残ってきました。
注:デニソワ人がネアンデルタール人の地方型として分岐したのは40万年前頃と解析されていますが、解剖学的現代
人類に組み込まれたネアンデルタール人の遺伝子はかなり後年の3-10万年前頃の集団からと考えられているよう
です。このため解剖学的現代人類に組み込まれたデニソワ人の遺伝子のほうが古い形態を残していたということ
になるようです。これは大事なことで、デニソワ型は地中海-中近東型に近いのではないかと考えられます。一
方、組み込まれたネアンデルタール人は終末期で、文化レベルも技術レベルも十分に成熟しきっていた、という
ことです。
注:記事「18-4. ホモサピエンスとは人類学上何者なのか」で触れたように、最新の研究で、ネアンデルタール人とデ
ニソワ人のY-DNAの変異型が同定され、恐らく80万年前頃と考えられるようになったY-DNA「Adam」に続き、
デニソワ人とネアンデルタール人が分化のツリー上に配置されるようになったのですが、「1-1. Y-DNAハプロタ
イプ 2019年6月版 ツリー」を参照ください、...。
未開の狩猟採集民でしかなかった原ホモサピエンスは、この成熟したネアンデルタール人と交雑することで一気に進化の爆発を起こし、 集団で行動するホモサピエンスの生活習慣を継承したハイブリッド種は恐らく人口繁殖力も極めて高く、圧倒的な人口増加の中に ネアンデルタール人を自然吸収する形で一体化し、解剖学的現代類に進化したと考えるのが最も合理的です。
3-3.解剖学的現代人類は原ホモサピエンスと終末期の成熟型ネアンデルタール人とのハイブリッド人類
現在のY-DNA情報から考える合理的な解剖学的現代人類の分化史は、
(1)出アフリカ後、中近東あたりで原ホモサピエンスと地中海-中近東型ネアンデルタール人が一次ハイブリッド。
Y-DNAはYAP型「DE」と非YAP型「CF」に分化。mtDNAはアジア型「M」とヨーロッパ型「N」に分化。
(2)「CF」はインド亜大陸〜中央アジア〜シベリアでデニソワ人と二次ハイブリッドし、「C」と「F」に分化し、 「C」は欧米の研究者によると「D」と行動を共にし、さらに東に向かって東遷したと考えられています。スンダ
ランドを進みサフール大陸に定着しオーストラロイドのニューギニア島人とオーストラリアンアボリジニとして
現代に残っています。また一部はスンダランドを北上しY-DNA「C」の子亜型群の中で最も古いY-DNA「C1a」が
日本列島に残っています。沖縄で発掘された縄文遺骨の復元顔はオーストラロイドの容貌です。「C」は現代のポ
リネシア人の主要子亜型でもあります。
(3)インド亜大陸に残った「F」は終末期のネアンデルタール人とハイブリッドした結果、更に「G」,「H」,「I」,
「J」,「K」と分化し、「G」,「I」,「J」はヨーロッパ大陸に向かい移動を開始したようです。「H」,「K」はイ
ンド亜大陸周辺にとどまったようです。「I」はクロマニヨン人から採取されたY-DNAでもあります。
(4)「K」はさらに分化を進め「K1」と「K2」に分化し、「K1」は更に「L」と「T」に分化しました。「L」はイン
ダス文明の主要遺伝子と推測され、現代のドラヴィダ人の主要遺伝子として残っています。「T」はアメリカ第3
代ジェファーソン大統領の遺伝子として有名です。
(5)「K2」からさらに「K2a」と「K2b」が分化し、「K2a」からさらに「N」,「O」が分化し東アジアの主役となり
ました。
「N」は遼河文明の主要亜型のようです。テュルク系の主要亜型として現代に残っています。またバルト3国の40
%を占める主要亜型でもあります。
「O」は中国国民の約40%、韓国国民の約70%、日本国民の約50%を占めている極東の最大亜型になります。日
本では長江系「O1」の弥生系水田稲作農耕民と黄河系「O2」の武装侵攻大王系や武士団系の主要亜型でもありま
す。中でも「O2」は出アジアし、東南アジアから、インド亜大陸やオセアニアに大展開し拡大をしています。
(6)残った子亜型「K2b」から孫亜型「K2b1」が分化し、出アジア後、太平洋に移動し「M」と「S」に分化しまし
た。
(7)最後に残った孫亜型「K2b2」から「Q」,「R」が分化し「Q」はシベリア経由で南北アメリカ大陸全土に拡散。
最後の最後の「R」の本体は西進しスラブ系「R1a」とケルト・ゲルマン系「R1b」としてヨーロッパの主役となり
ました。
以上のことから、出アフリカしたすべての現代人類はネアンデルタール人の遺伝子を持っており、特にY-DNA「C」の集団はデニソワ人の 遺伝子も受け継いでいる分析結果になったものと考えられます。
以上が、現在の全世界のY-DNA分化ツリーから最も考えられうるストーリーです。当ガラパゴス史観が生きているうちに 全容解明することを期待しましょう。
西欧列強が世界中を植民地化するべく搾取活動を続けているときにわかったことは、アフリカ大陸やニューギニア・オーストラリアや アンダマン島の先住民は、何万年もの間、狩猟採集民の文化レベルにとどまっていた、ということでした。研究調査からかなり高度な 文化・技術レベルに達していたと判ってきているネアンデルタール人と比べると、分類学的にホモサピエンスに進化したというだけでは 現代文明には到達できないという証明でしょう。何かブレークスルーのファクターがあったと考えるべきでしょう。
出アフリカ後に、古いネアンデルタール人とハイブリッドしたというだけでは古ホモサピエンスのレベルで まだ解剖学的現代人類には至らず、インド亜大陸−中央アジア周辺で、成熟した文化を築き上げていた終末期のネアンデルタール人と ハイブリッドしたことで初めて現代人類として完成(つまり解剖学的現代人類)したのではないかと考えるのが最も合理的です。
3-4.解剖学的現代人類は進化の爆発に遭遇できたのではないか!!!
終末期ネアンデルタール人と出アフリカ組古ホモサピエンスの交雑の結果、進化の爆発が起きたと推測するのが最も妥当です。
前述のようにホモサピエンスは華奢型の突然変異型だったため、生き残るために集団化したことで高い人口増加率(繁殖力)を得たのに対し、 家族単位のため比較して少ない人口だったネアンデルタール人がハイブリッドしながら自然吸収される形で統合されたのが 解剖学的現代人類=ハイブリッド型と考えるのが最も妥当性が高いのです。
つまり解剖学的現代人類とは、先進文化を持っていた頑丈型の終末期ネアンデルタール人と華奢型の狩猟採集民だった 古ホモサピエンスとのハイブリッドの結果誕生し、進化の爆発に遭遇できた人類と考えるのが最も妥当でしょう。
ホモサピエンスがもし出アフリカせずネアンデルタール人とも出会わずアフリカの中に留まっていたら、 人類は相変わらず19世紀ごろのサン族やピグミー族のように素朴な狩猟採集段階に留まっているだろうと容易に推測できますが、 北京原人やジャワ原人などのホモエレクトスも出アフリカし、ネアンデルタール人も出アフリカしたということは、 原ホモサピエンスが出アフリカしたのは人類の遺伝子が導く宿命ではないかとも思われます。
それでも出アフリカした原ホモサピエンスが、ネアンデルタール人と交配しハイブリッド型に進化し狩猟採集文化から脱し、 現代文明にまで至ったことまで必然だったかというと、それは今後の研究待ちですね。
4.日本列島への最初の到来者は、古代亜型集団:Y-DNA「D」と「C」
4-1.縄文系遺伝子集団:Y-DNA「D」の移動
さてY-DNA「D」は、欧米の研究者の考えではY-DNA「C」と共に東遷したと欧米の研究者は考えられており、現代より120m〜140mも 海面が低かったために陸地だったと考えられるインド亜大陸沿岸の大陸棚に沿って東進しインド亜大陸をスルーしスンダランドに到達し、 そこから北上し現在の中国大陸に到達したようです。しがってY-DNA「D」は「C」とともに終末期の成熟期のネアンデルタール人と ハイブリッドしておらず、進化の爆発に遭遇できなかった古ホモサピエンスの段階に留まっていると考えるのが合理的です。 その時に大陸棚だった現在のアンダマン諸島域に定住したY-DNA「D」集団は、その後の海面上昇で島嶼化した現アンダマン諸島で 孤立化し現代までJarawa族やOnge族などの絶滅危惧部族として古代亜型Y-DNA「D」を伝えてきています。 研究者によっては中国大陸−インド亜大陸経由で南下してきた、とする説もありますが、納得には至りません。
このためY-DNA「D」は基本的に古代性の強い狩猟採集民と考えてよく、日本人の持つ古代的なホスピタリティの源泉であることは 間違いないです。
以下の写真は記事「30-11. 縄文遺伝子Y-DNA「D2」」でご紹介したものです。残念ながら現在Wikipediaでは写真は削除されていました。

Y-DNA「D」が100%の純系子孫のOnge族です。白黒写真なのでわかりませんが Onge族やJarawa族は世界で最も黒い(ネイティヴアフリカンより黒い)と言われているそうです。Y-DNA「D」が日本列島に渡って来たのは、 オーストラリアのアボリジニは5−6万年前頃には既にサフール大陸に到達していたらしいので、日本列島にも恐らく遅くても 3−4年前頃には渡来していたと考えるのが妥当でしょう。その当時のY-DNA「D」集団は黒かったのか既に黄色くなっていたのかは 全く情報がありません。しかし確かなのは鯨面文身であったことでしょう。Y-DNA「D」が85%を占める子孫のアイヌが鯨面文身だったのは、 子孫として数万年にわたり正しく文化を残してきたことになります。
Jarawa族やOnge族は現代でもネグリートなので、当然「D」系縄文人も小柄なネグリートつまり、「倭」人であったことは間違いないでしょう。
その時にY-DNA「C」集団の本体は更に東進しサフール大陸に到達したようです。このY-DNA「D」と「C」の移動は、 Y-DNA「C」が習得した沿岸渡海技術で思いのほか早く5万年前にはサフール大陸に到達したと推測できます。
4-2.縄文系ハンター遺伝子集団:Y-DNA「C」の移動
Y-DNA「CT」から分離したもう一方の移行亜型Y-DNA「CF」は恐らくインド亜大陸到達までに古代亜型Y-DNA「C」とY-DNA「F」に分離し、 Y-DNA「F」はインド亜大陸に留まりそこで再度先住ネアンデルタール人と交雑した結果、Y-DNA「G」以降の全ての現代Y-DNA亜型の 親遺伝子となったと推測できます。こうしてインド亜大陸は現代Y-DNA亜型全ての発祥の地となったと考えるのが今のところ合理的です。
もう一方の分離した古代亜型Y-DNA「C」は、欧米の研究者の説明ではY-DNA「D」と行動を共にしたらしく東進しスンダランドに入り、 一部はY-DNA「D」と共に中国大陸に到達し、本体はそのまま更に東進しサフール大陸に到達したと考えられています。 サフール大陸に入った集団はサフール大陸内で拡大し、海面上昇後分離したニューギニアとオーストラリア大陸にそれぞれTehit族、 Lani族やDani族などニューギニアの先住民集団やオーストラリア・アボリジニ集団、つまり共にオーストラロイドとして現代まで残っています。
そして5万年前にはオーストラリアに到達していた集団の遺跡から回遊魚のマグロの骨が東海大学らの調査により発見され、 Y-DNA「C」は沿岸を船で移動できる海洋性ハンター集団だったと考えられます。 従ってサフール大陸に到達したY-DNA「C」集団は更にそのまま船で海に漕ぎ出し、ポリネシア全土に拡大していったようです。 ポリネシアのY-DNAの主要亜型として検出されるY-DNA「C」は、実は縄文の海洋性ハンターY-DNA「C」と同じ亜型です。 つまり日本列島で検出される海洋性ハンター遺伝子Y-DNA「C」亜型は、現代ポリネシア人と同じ先祖を持つことになります。
ところがオセアニアの遺跡の年代はそれほど古くはないようです。海面上昇で当時の陸地は全て水没し遺跡も当然海中です。 海中考古学が進歩すれば、Y-DNA「C」集団がポリネシアに漕ぎ出したのはかなり古いという事が判るのではないかと期待しています。
注:一部の日本人の持つ海洋性気質は、このポリネシア人と共通の祖先Y-DNA「C」から受け継いだ気質と言っても差
し支えないでしょう。
前述のように縄文時代を形成したと思われるY-DNA「D」と「C」は、古代亜型としてネアンデルタール人と一次交雑を経験したと 考えられますが、古ホモサピエンスの段階に留まっており、解剖学的現代人類=現ホモサピエンスの段階には至っていなかったと考えられます。 弥生時代以降に後に新しい現代亜型と交雑することで現代性を身に着けることができたのではないかと推測できます。 これは極めて大胆な推測ですが、これに関する知見、否定する知見はまだ全く報告されていません。
以下の写真は記事「30-12. 縄文ハンター遺伝子Y-DNA「C1a」,「C3a」」でご紹介したものです。残念ながら現在Wikipediaでは 写真は削除されていました。

Y-DNA「C」が100%の純系子孫のニューギニアのLani族です。濃い褐色で彫深のソース系になります。Y-DNA「D」のJarawa族やOnge族同様ネグリートです。 出アフリカ時のホモサピエンスはY-DNA「B」と考えられ、「B」は現在ピグミー集団として知られています。 ということは、恐らく古代亜型遺伝子のY-DNA「D」,「E」,「C」,「F」は全てネグリートだったと思われます。 日本人は近代になっても男一匹五尺の体、と言われていた通り、日本人の男性人口の約50%を占める縄文系Y-DNA「D」と「C」は、 戦前までネグリート形質を強固に維持してきたのです。戦後の食糧事情や衛生環境の圧倒的な改善で日本人の身長は伸びてきましたが、 記事「19-14. 縄文遺伝子Y-DNA「D2」のネグリート(倭人)体質」でご報告した通り、戦後一気に伸びた身長も男女ともに1980年代でピタッと止まりました。 女性の縄文系の割合はほぼ67%です。ネグリート形質は強固で、食糧事情や衛生環境の圧倒的な改善にも打ち勝ち、日本人の身長を抑え込んでいるようです。 イタリアやスペインなどのラテン系の人々の低い身長もY-DNA「E」の持つネグリート形質の特徴でしょう。 欧米人の高身長の源は、ヨーロッパ中に拡散した北欧系の高身長遺伝子によるものです。
というわけでY-DNA「C」もやはり鯨面ですが、文身はしていないようです。Y-DNA「D」とはかなり異なる集団だということがわかると思います 。同じY-DNA「C」のアボリジニの祖先は沿岸でマグロのような回遊魚を漁していた渡海技術を持つ技術集団であることが判っています。 このY-DNA「C」集団は一時はニューギニア全島に分布していたと思われますが、かなり後代にY-DNA「S」と「M」のメラネシア人が 移動してきたときに押し出されたと思われ、ポリネシア全域に拡大し現在のポリネシア人のコア遺伝子となっていったと考えられます。 渡海技術を持っていたため、出ニューギニアできたようです。Lani族やDani族、Tehit族等純系部族は取り残されたのか、 やや内陸に居住しているので内陸適応したのかもしれません。
4-3.Y-DNA「D」の先輩チベット人の高高地適応獲得形質:
欧米の研究者は、標高の高い高地に適応しているチベット人の高高地適応獲得形質は、デニソワ人との交雑で得た後天的獲得形質と 考えているようです。
2019年度のノーベル生理学賞「細胞が低酸素を検知し応答する仕組みの発見」では、低酸素状態になると「エリスロポエチンが増えて 赤血球が多く作られ酸素を確保する」仕組みだそうですが、負の問題として赤血球が増えるとドロドロ血になり、高地では心臓への負担が 大きくなるという高血圧症や新生児の低体重、死亡の原因になるのだそうです。特に妊婦さんの負担は極めて大きいのだそうです。
その血液に酸素を供給するヘモグロビンの生成を調節するのがEPAS1遺伝子だそうです。チベット人は高地になってもヘモグロビンの 過剰生成を抑えられるように変異しており、標高4000mを越える高地でも低酸素症の発症を抑えることができているのだそうです。 このチベット人のEPAS1変異が、解読されたデニソワ人のEPAS1の変異と同じだったのだそうです。この研究結果は2014年にNature に 発表されていました。
ところがニューギニア高地人を含むメラネシア人にはこの遺伝子変異が受け継がれていないのだそうです。EPAS1遺伝子とは 別の作用機序があるのかもしれません。アンデスのマチュピチュなど先住民文化の子孫の人々はY-DNA「Q」ですが、 まだEPAS1遺伝子の報告はないようです。
4-4.Y-DNA「D」の弟亜型のY-DNA「E」とは?
さて戻って、移行亜型Y-DNA「DE」はさらに古代遺伝子Y-DNA「D」とY-DNA「E」に分化しましたが、Y-DNA「D」がインド洋沿岸に沿って 東進したのに対し、Y-DNA「E」は逆に西進し地中海南北沿岸に定着し、地中海南岸(アフリカ北岸)に移動した集団はさらにアフリカ全土に 展開し、始祖亜型である原ホモサピエンスの先住民の中に入り込み、現代では、始祖亜型Y-DNA「A」と「B」のネイティヴ・アフリカン集団の 中に古代亜型Y-DNA「E」の遺伝子が混在するようになっています。
注:現ネイティヴ・アフリカンのコイサン集団にもピグミー集団にも、Y-DNA「A」と「B」の純系部族は存在していま
せん。このため原ホモサピエンスのイメージは推測の範囲を出ません。
注:アフリカ大陸にはその後Y-DNA「R1a」と分化したY-DNA「R1b」(後代のケルト系遺伝子)集団がアナトリア、
中近東から南下してきて、更に新しい集団として現在のカメルーンあたりを中心にネイティブ・アフリカンの一部
を形成しています。
しかし出戻りアフリカした古代亜型Y-DNA「E」は古ホモサピエンスの段階で、進化の爆発が進む前にアフリカ大陸に入ってしまったため、 基本的に狩猟採集のままとどまり、ユーラシア大陸で起きた農耕革命など進化の爆発に会わないまま現代に至っています。
ところが地中海北岸に定着したY-DNA「E」は、その後ヨーロッパに移動してきたY-DNA「G」,「I 」,「R」などの現代亜型と 刺激しあいながら集団エネルギーを高め、ローマ帝国やカルタゴなどの文明を築くまでに至りました。要するにアフリカ大陸のような 原ホモサピエンスの始祖亜型との遭遇では同化してしまい、文明を興すような爆発的進化は起こりませんでしたが、 ヨーロッパ大陸ではより新しい現代亜型との遭遇が集団エネルギーを高める要因になったのでしょう。
4-5.縄文人のコアをなすY-DNA「D1b」,「C1a」の日本列島への上陸
最初の縄文人はY-DNA「D1b」(旧「D2」)を多数派とする、Y-DNA「C1a1」(旧C1a」との混成部隊でしょう。
スンダランドから北上し現在の中国大陸に入ったY-DNA「D」とY-DNA「C」の混成集団は中国大陸の先住集団として拡大したはずです。 この時に混成集団の一部の集団は中国大陸には入らずにさらに北上し、当時海面低下で大きな川程度だった琉球列島を渡ったと思われます。 集団はそのまま北上し現在の九州に入った可能性が大。また一部は日本海の沿岸を北上し当時陸続きだったサハリンから南下し北海道に入り、 当時同様に川程度だった津軽海峡を渡り本州に入った可能性も大です。つまり日本本土への入り方が2回路あった可能性が大なのです。
現在沖縄・港川で発掘される遺骨から復元再現される顔は完璧にオーストラロイドの顔です。と言うことは、スンダランドから 北上の途中、沖縄に定住した「D」と「C」の混成集団がその後の琉球列島人の母体になり、 サハリンから南下した「D」集団がのちのアイヌ人の集団になった可能性が極めて大と推測できます。
さて中国大陸に展開したY-DNA「D」は残念ながら後発のY-DNA「O」に中国大陸の中原のような居住適地から駆逐され、 南西の高地に逃れY-DNA「D1a」のチベット人や羌族の母体となったようです。そして呪術性が高い四川文明は、Y-DNA「C」と「D」が残した 文明と考えられます。このため同じY-DNA「C」と「D」遺伝子を合わせて50%弱も持つ日本人には、四川文明の遺物は極めて親近感が あるのかもしれません。
日本の民話とチベットの民話には共通性がかなりありますが、これらはY-DNA「D」が伝えてきた民話と考えて差し支えないでしょう。
しかし一緒に移動したと考えられる-DNA「C」の痕跡は現在の遺伝子調査ではチベット周辺では検出されていません。 どうやら途絶えてしまった可能性が高い、もともと海洋性の遺伝子なので、内陸の高地は居住適地ではなかったのかもしれません。 縄文時代に火炎土器のような呪術性の強い土器を製作したと考えられるY-DNA「C」なので、四川文明の独特な遺物類はY-DNA「C」が製作した 可能性が極めて高いのですが、Y-DNA「D」のようにデニソワ人からチベット高原のような高高地適応形質を受け継がなかったため 途絶えてしまった可能性も考えられますね。チベット人の高高地適応がデニソワ人から受け継いだということは、 四川文明もデニソワ人が加わっていた可能性はあります、Y-DNA「C」とデニソワ系Y-DNA「D」との合作かもしれません。
スンダランドから琉球列島を北上した集団(Y-DMA「D1b」とY-DNA「C1a」は、一部は琉球列島に留まり、琉球人の母体となったのでしょう。 しかし、そのまま更に北上し九州に到達したかどうかはまだ推測できていなませんが、日本各地に残る捕鯨基地や水軍など 日本に残る海の文化は海洋性ハンターと考えられるY-DNA「C1a」がそのまま北上し本土に入った結果と考えるのが最も合理的です。
前述のようにオーストラリアの海洋調査で、数万年前にY-DNA「C」の時代にすでに漁労が行われ、回遊魚のマグロ漁が行われていたと 考えられる結果のマグロの魚骨の発掘が行われ、Y-DNA」「C」はスンダランドからサフール大陸に渡海する手段を持ち、更に漁をする レベルの船を操る海の民であったことが考えられています。このことはスンダランドから大きな川程度だった海峡の対岸の琉球列島に 入ることはさほど困難ではなかったと考えられ、Y-DNA「C」と交雑し行動を共にしていたと考えられるY-DNA「D」も一緒にさらに北上し 本土に入ったことは十分に考えられます。すべての決め手はネアンデルタール人から受け継いだと考えられるY-DNA「C」の 海洋性技術力のたまものでしょう。
一方日本海をさらに北上したY-DNA「D」集団があったことも十分に考えられます。この集団はサハリンから南下し北海道に入り、 更に大きな川程度だった津軽海峡を南下し、本土に入ったと考えられ、サハリンや北海道に留まった集団は古代原アイヌ人の母体と なったでしょう。この時Y-DNA「C1a」は北海道には留まらず恐らく本州北部の漁民の母体となり、Y-DNA「D1b」は蝦夷の母体となったでしょう。
このY-DNA「D1b」とY-DNA「C1a」が縄文人の母体と言って差し支えないでしょう。つまり縄文人は主力の素朴な狩猟採集集団の Y-DNA「D1b」と渡海技術や土器作成などの技術力を持つ海洋性ハンターのY-DNA「C1a」との混成集団であると推測できます。 この海洋性ハンター遺伝子が一部日本人の持つ海洋性気質の源流でしょう。日本人は単純な農耕民族ではないのです。
4-6.さらにY-DNA「C2a」の日本列島への上陸
ところがサハリンから南下せずにシベリア大陸に留まり陸のハンターに転身したのが大陸性ハンターY-DNA「C2」(旧「C3」)でしょう。 この集団はクジラの代わりにマンモスやナウマンゾウを狩猟する大型獣狩猟集団に変身したと思われます。 ところが不幸にもシベリア大陸の寒冷化によりマンモスもナウマン象も他の大型獣も少なくなり移住を決意したのでしょう。 一部はナウマン象を追って南下し対馬海峡を渡り本土に入りY-DNA「C2a」(旧C3a」)となり山の民の母体となったでしょう。 また一部はサハリンからナウマンゾウの南下を追って北海道、更に本土へ渡った集団もあったでしょう、 北の山の民の母体となったと推測できます。
この山の民になった大陸性ハンターY-DNA「C2a」が縄文人の3つ目の母体と考えてよいでしょう。つまり縄文人とは、 核になる狩猟採集民のY-DNA「D1b」と海の民のY-DNA「C1a」及び山の民のY-DNA「C2a」の3種混成集団と考えられます。
このY-DNA「C2a」が一部日本人の持つ大陸性気質の源流と考えられるでしょう。Y-DNA「C1a」は貝文土器など沿岸性縄文土器の製作者、 Y-DNA「C2a」は火炎土器など呪術性土器の製作者ではないかと推測され、いずれにせよ縄文土器は土器作成の技術を持つ Y-DNA「C」集団の製作と推測され、Y-DNA「D」は素朴な狩猟採集民だったと推測できます。
この山の民のY-DNA「C2a」が南下するときに、南下せずY-DNA「Q」と共に出シベリアしたのがY-DNA「C2b」(旧「C3b」)の一部であろう と考えられます。このY-DNA「Q」はヨーロッパでは後代のフン族として確定されています。このY-DNA「Q」はシベリア大陸を横断するような 移動性の強い集団だったようです。シベリア大陸を西進せずに東進し海面低下で陸続きになっていたアリューシャン列島を横断し 北アメリカ大陸に到達し、Y-DNA「Q」が更に南北アメリカ大陸全土に拡散したのに対し、Y-DNA「C2b」は北アメリカ大陸に留まり ネイティヴ・アメリカンの一部として現代に遺伝子を残しています。最も頻度が高いのはTanana族で現代でも約40%もの「C」頻度を保持しています。 北アメリカや中米で発掘される縄文土器似の土器の製作者は当然土器作成技術を持っていたこのY-DNA「C2a」ではないかと推測できます。
またそのままシベリア大陸/東北アジアに留まったY-DNA「C2」はさらに孫亜型Y-DNA「C2b1a2」に分化し、大部分はモンゴル族や ツングース族の母体となりました。また一部だった古代ニヴフ族は北海道に侵攻しY-DNA「D1b」の原アイヌ人を征服しオホーツク文化を 立ち上げたと考えられます。本来素朴な狩猟採集民だった原アイヌ人は支配者の古代ニヴフの持つ熊祭りなどの北方文化に変化し、 顔つきも丸っこいジャガイモ顔からやや彫の深い細長い顔に変化したようです。明治時代に欧米人からコーカソイドではないかと言われた 一部のアイヌ人の欧米顔はオホーツク文化人の名残でしょう、原アイヌ人の風貌ではありません。現代アイヌ人の持つ風習から北方性の 風俗・習慣を除くとほんの少し残っているかもしれない原アイヌ人=縄文人の文化が構築できるかもしれないです。
5.長江文明系稲作農耕文化民の日本列島到来
さて、日本人は農耕民族と言われますが、果たしてそうなのでしょうか?縄文人は明らかに農耕民族ではありません。 狩猟採集民とハンターの集団だったと考えられます。ではいつ農耕民に変貌したのでしょうか?
前述のように、古代遺伝子Y-DNA「F」から分化した現代遺伝子亜型群はY-DNA「G」さらに「H」、「I」、「J」、「K」と分化し、 Y-DNA「K」からY-DNA「LT」とY-DNA「K2」が分化しました。 このY-DNA「LT」から更にY-DNA「L」が分化しインダス文明を興し、後にドラヴィダ民族の母体となったと考えられています。 Y-DNA「T」からは後のジェファーソン大統領が出自しています。
Y-DNA「K2」はさらにY-DNA「K2a(NO)」とY-DNA「K2b」に分化し、Y-DNA「NO」が更にY-DNA「N」とY-DNA「O」に分化しました。 このY-DNA「N」は中国の遼河文明を興したと考えられているらしいのです。 このY-DNA「N」は現在古住シベリア集団(ヤクート人等)に濃く70-80%も残されており、テュルク(トルコ)系民族の母体と考えられています。
しかし現代トルコ人は今のアナトリアに到達する過程で多種のY-DNAと混血し主力の遺伝子は Y-DNA「R1a」,「R1b」,「J2」などに変貌している為、東アジア起源の面影は全くありません。 唯一タタール人に最近まで若干の面影が残っていましたが、今のタタール人もY-DNA「R1a」が主力に変貌してしまっています。 Y-DNA「N」はシベリア大陸の東西に高頻度で残りバルト3国の主力Y-DNAとして現代も40%以上も残っています。 やはり移動性の強い遺伝子のようです。
さていよいよ日本農耕の起源に触れなければなりません。Y-DNA「NO」から分化したもう一方のY-DNA「O」は、 中国の古代遺跡の発掘で、古代の中国大陸居住人は現在のフラットな顔つきと異なり、コーカソイドの面影が強いと報告されている事は研究者の周知である。 つまり本来の現代亜型人類は彫が深かったといってよく、現代東北アジア人のフラット/一重まぶた顔は 寒冷地適応に黄砂適応が加わった二重適応の特異的な後天的獲得形質と言って差し支えないでしょう。 (余談ですが、人類(動物)は体毛が減少する方向に進んでいるので、実は禿頭/ハゲ頭も「進化形態」であると言えます。)
この東北アジア起源のY-DNA「O」は雑穀栽培をしていたようです。東アジア全体に拡散をしていったようです。 日本列島では極低頻度ですがY-DNA「O」が検出されています。陸稲を持ち込んだ集団と考えられます。 東北アジアの住居は地べた直接だっと考えられます。主力集団は黄河流域に居住していたため、 長年の黄砂の負荷で現代東アジア人に極めてきついフラット顔を獲得したのでしょう。
一方南下し温暖な長江流域に居住した集団から長江文明の稲作農耕/高床住居を興したY-DNA「O1a」と「O1b」が分化し、 更にY-DNA「O1b1」(旧「O2a」)と「O1b2」(旧「O2b」)が分化し水田稲作農耕は発展したようです。 現在の分布地域から推測すると、Y-DNA「O1a」は楚民、Y-DNA「O1b1」は越民、「O1b2」は呉民の母体と推測できます。
5-1.長江文明系稲作農耕文化民の拡散
長江文明は黄河文明に敗れ南北にチリジリになり、Y-DNA「O1b1」の越民は南下し江南から更にベトナムへ南下し、 更に西進しインド亜大陸に入り込み農耕民として現在まで生き残っています。 ほぼ純系のY-DNA「O1b1」が残っているのはニコバル諸島(Y-DNA「D*」が残るアンダマン諸島の南に続く島嶼でスマトラ島の北に位置する) のShompen族で、Y-DNA「旧O2a」100%の頻度です。
WikipediaのShompen族の写真

真ん中の人物は近藤正臣かキムタクか、日本に普通にいる風貌ですね。稲作農耕民の典型と考えてよいでしょう。
また南インドのドラヴィダ民族中には検出頻度がほとんどY-DNA「O1b1」のみの部族もあり、 越民がいかに遠くまで農耕適地を求めて移動していったか良く分かります。 カースト制度でモンゴロイドは下位のカーストのため、他の遺伝子と交雑せず純系の遺伝子が守られてきたようです。 この稲作農耕文化集団である越民の子孫のドラヴィダ民族内移住が、ドラヴィダ民族(特にタミール人)に 長江文明起源の稲作農耕の「語彙」を極めて強く残す結果となり、その結果、学習院大学の大野教授が 日本語タミール語起源説を唱える大間違いを犯す要因となりましたが、 こんな遠くまで稲作農耕民が逃げてきたことを間接証明した功績は極めて大きいのです。
5-2.日本列島と朝鮮半島の稲作農耕民の共通点
一方、呉民の母体と考えられるY-DNA「O1b2」は満州あたりまで逃れ定住しましたが、更に稲作農耕適地を求め南下し朝鮮半島に入り定住し、 更に日本列島にボートピープルとして到達し、先住縄文人と共存交雑しY-DNA「O1b2a1a1」に分化したと考えられます。 この稲作農耕遺伝子Y-DNA「O1b2」は満州族で14%、中国の朝鮮族自治区で35%、韓国で30%、日本列島でも30%を占めます。 この満州族の14%は、満州族の中に残る朝鮮族起源の姓氏が相当あることからやはり朝鮮族起源と考えられ、 呉系稲作農耕文化遺伝子を現在に残しているのは朝鮮民族と日本民族のみと断定して差し支えないでしょう。 この共通起源の呉系稲作農耕文化の遺伝子が日本人と朝鮮人の極めて近い(恐らく起源は同一集団)要因となっています。 北朝鮮はツングース系遺伝子の分布が濃いのではないかと考えられますが、呉系の遺伝子も当然30%近くはあるはずです。 過去の箕子朝鮮や衛氏朝鮮が朝鮮族の起源かどうかは分かりませんが、呉系稲作農耕民が朝鮮族の起源の一つであることは間違いないでしょう。
6.黄河文明系武装侵攻集団の到来
狩猟採集と海陸両ハンターの3系統の縄文人と、長江系稲作農耕文化の弥生人が共存していたところに、 武装侵攻者として朝鮮半島での中国王朝出先機関内の権力闘争の戦いに敗れ逃れてきたのが、 Y-DNA「O2」(旧「O3」)を主力とする黄河文明系集団でしょう。 朝鮮半島は中華王朝の征服出先機関となっており、 長江文明系とツングース系が居住していた朝鮮半島を黄河系が占拠して出先機関の「郡」を設置し、 韓国の歴史学者が朝鮮半島は歴史上だけでも1000回にも及び中華王朝に侵略された、と言っている結果、 現代韓国は43%以上のY-DNA「O2」遺伝子頻度を持つ黄河文明系遺伝子地域に変貌してしまいました。
朝鮮半島での権力闘争の戦いに敗れ追い出される形で日本列島に逃れてきた集団は、当然武装集団でした。 おとなしい縄文系や和を尊ぶ弥生系を蹴散らし征服していったようです。長江系稲作農耕集団は、 中国本土で黄河系に中原から追い出され流れついた先の日本列島でも、また黄河系に征服されるという二重の苦難に遭遇したようです。
この黄河系集団は日本書紀や古事記に言う天孫族として君臨し、その中で権力闘争に勝利した集団が大王系として確立されていったようです。 この黄河系武装集団の中に朝鮮半島で中華王朝出先機関に組み込まれていた戦闘要員としてのツングース系の集団があり、 ともに日本列島に移動してきた可能性が高いY-DNA「P」やY-DNA「N」でしょう。 好戦的な武士団族も当然黄河系Y-DNA「O2」でしょう。出自は様々で高句麗系、新羅系、百済系、渤海系など 朝鮮半島の滅亡国家から逃げてきた騎馬を好む好戦的な集団と推測できます。
この黄河文明系Y-DNA「O2」系は日本列島で20%程度検出される重要なY-DNAです。韓国では43%にもなり、 いかに黄河文明=中国王朝の朝鮮半島の侵略がひどかったが容易に推測できます。 日本列島の長江文明系Y-DNA「O1b2」系と黄河系Y-DNA「O2」系は合計50%近くになります。韓国では73%近くになります。 つまり日本人の約50%は韓国人と同じ長江文明系+黄河文明系遺伝子を持つのです。これが日本人と韓国人が極めて似ている理由であります。
一方、韓国には日本人の約50%を占める縄文系Y-DNA「D1b」,Y-DNA「C1a」とY-DNA「C2a」が欠如しています。 これらY-DNA「D1b」,「C1a」とY-DNA「C2a」は日本人の持つ素朴なホスピタリティと従順性と調和性の源流であり、 このことが日本人と韓国人の全く異なる民族性の大きな理由であり、日本人と韓国人の近くて遠い最大の要因になっています。
一方、日本人の持つ一面である残虐性/競争性/自己中性等は、20%も占める黄河系Y-DNA「O2」系からもたらされる 特有の征服癖特質が遠因と言って差し支えないような気がします。
7.日本人の源流....簡易まとめ
日本人の持つ黙々と働き温和なホスピタリティや和をもって貴しとする一面と、 一方過去の武士団や維新前後の武士や軍人の示した残虐性を持つ2面性は、 日本人を構成するもともとの遺伝子が受けてきた歴史的な影響の結果と言えそうです。
日本人の3つの源流は、
・日本列島の中で約1万年以上純粋培養されてきた大多数の素朴な狩猟採集民と少数のハンターの縄文系、
・中国大陸から僻地の日本列島にたどり着き、集団の和で結束する水田稲作農耕民の弥生系、
・朝鮮半島から追い出されてきた、征服欲出世欲旺盛な大王系/武士団系の武装侵攻集団系、
個人の性格の問題では解説しきれない、遺伝子が持つ特質が日本人の行動・考えに強く影響していると思えます。 世界の技術の最先端の一翼を担っている先進国で、50%もの古代遺伝子(縄文系、しかも女系遺伝するmtDNAでは何と約67%が 縄文系のmtDNA「M」系なのです。)が国民を構成しているのは日本だけで極めて異例です。 もしこの縄文系遺伝子がなければ、日本列島と朝鮮半島及び中国はほとんど同一の遺伝子圏/文化圏と言って差し支えないでしょう。 それだけ縄文系遺伝子がもたらした日本列島の基層精神文化は、 日本人にとって世界に冠たる独特の国民性を支える、守るべき大切な資産なのです。
8.Y-DNAの分化.....エピジェネティクスか
これまで独立した亜型として扱われてきた近代亜型のY-DNA「L」,「M」,「N」,「O」,「P」,「Q」,「R」,「S」,「T」は、 現在、再び統合されてY-DNA「K」の子亜型Y-DNA「K1」とY-DNA「K2」の更に子亜型(孫亜型)として再分類される模様です。 つまり独立名をつける亜型群として扱うほど「違いが無い」ということなのです。
ところがこのY-DNA「K」は、我々極東の代表Y-DNA「O」や西欧の代表Y-DNA「R」や南北ネイティヴアメリカンの Y-DNA「Q」等が含まれているのです。とても遺伝子が近いとは思えないのです。では何故これほど外観も行動様式も異なるのでしょうか?
これらの亜型群は、何十万年の歴史でユーラシア大陸の各地で亜種に近いほど分化していたと考えられている ネアンデルタール人やデニソワ人のY-DNA亜型を受け継いだだけの可能性も十分にあるのです。 西欧と極東であまりにも異なる外観や行動様式などの違いの原因をネアンデルタール人との接触に求めるのは荒唐無稽とは言えないでしょう。 我々現代人は、当然ですが、各亜型群が分化した当時の形態は全く知りませんが、各亜型に分化してから各地に移動したのか、 移動した先の地域に順応して分化したのか、いずれかでしょう。 分化の要因はまだ推測できていませんが、移住先の地方型先住ネアンデルタール人との交雑、気候や食生活の変化、 移住先に特有のRNAウイルスとの共生化.......等々、いずれにせよ後天的な獲得形質ではないかと思われます。
ネアンデルタール人からホモサピエンスが分化したのは、骨形成不全、筋肉形成不全等、身体能力の発現不全症だった可能性も高いのです。 つまりひ弱な華奢型の退行型突然変異だったような気がしますが、 Y-DNAの亜型の分化は突然変異というよりエピジェネティクス(後天的な獲得形質)と考えるほうが理にかなっていると思います。
9.雑考
(極めて余談ですが、北方系極東人の多くは寒冷地適応や黄砂適応を受け、フラット顔になってはいますが、 中国で発掘される古代人骨はほとんどコーカソイド顔であり、フラット顔は後天的獲得形質であることは研究者達が認めています。 日本人にはこの後天的獲得形質を獲得してから日本列島に渡ってきた集団が多かったことを示しています。 日本人の胴長短足は、高身長の弥生系と武装侵攻系の上半身と小柄な縄文の下半身の交雑の結果に過ぎず、日本人に意外に多い反っ歯や受け口も 弥生系の細身の顎に縄文系のがっしり歯列が収まりきらず前に出てしまっただけであり、親知らずは逆に出られなかっただけなのでしょう。
また日本人固有の古代的なホスピタリティは、縄文系である古代亜型Y-DNA「D」と「C」(合計で日本人男性の出現頻度約45%を占める) 及びmtDNA「M」(合計で日本人全体の約67%を占める)の(古ホモサピエンス)固有の特質であり、現代亜型群の特質ではなさそうです。 つまり特に日本人と他の民族との違いのほとんどは、この縄文系遺伝子の伝えてきた極めて古代的な、 狩猟採集民やハンター民の持つ行動様式や思考回路のもたらす結果に帰することは疑いようがありません。
もし、天孫族や武士団族が朝鮮半島から負け組として渡来してこなければ、日本列島は徳川時代の高度な文化もなく、 当時世界最大の都市だった江戸もなく、容易に西欧列強の植民地になっていたでしょう。つまり極めて残念なことですが、 日本人の世界に冠たる高度な技術力や文化性は、日本列島の3重遺伝子構造を構成する遺伝子の中で最後にやってきた黄河系 Y-DNA「O2」(旧O3)が牽引してきたものと言って間違いないでしょう、中国や韓国と支配階級が同じY-DNA「O2」遺伝子なのに結果が異なってきたのは、 中韓が常に外敵との抗争や侵略に脅かされ、技術や文化の熟成が現代に入るまでまで確立出来なかったのに対し、 日本列島は江戸時代には出来ていたという違いなのでしょう。
10.時代の趨勢
記事「3-3. Y-DNA「R1b」 ケルト度 調査」に書いた文章を復誦します。
極めて明らかなことは、国・国民が先進的になるには純系民族では無理なのです。辺境民化してしまいエネルギーが低すぎるのですが、 競う共存遺伝子の種類が多ければ多いほど集団エネルギーが高くなり、国の活性度が上がり、覇権に向かうのです。 アジアの中で唯一近代化に成功した日本は縄文系−弥生系(長江文明系)−武装侵攻系(黄河文明系)が交雑し、 武装侵攻系が核になり集団エネルギーを一気に高め、一時はジャパンアズNo1と覇権を握るかもしれないほどの勢いを手に入れました。
しかし日本が高止まりしてしまった間に中国が、日本以上の複雑な遺伝子ミックスにより集団エネルギーを高め、近代化らしきものに成功し 対外的には日本に取って替わりアジアの覇権を握ったように見えるレベルに達しました。しかし国民一人当たりの生産性があまりにも低く、 日本の1/4以下程度しかなく真の覇者には恐らく永久になれないでしょう。
その中国も恐らく近いうちに日本同様高止まりするでしょうが、東アジアには日本、中国に代わる国はもはや存在しません。 南アジアのインドはロシアのスラブ系と同じインド・ヨーロッパ系遺伝子が支配する国で複雑な遺伝子ミックをもっていますが、 中国同様あまりに国民一人あたりの生産性が低すぎ、日本の1/20程度しかなく覇権には届かないでしょう。
当分の間はY-DNA「R1b」のアメリカとY-DNA「O2」(旧「O3」)の中国が覇権争いを続けるでしょうが、中国も日本もアメリカに 対する輸出で生産性を上げてきたので、アメリカにとって代わることは逆に自滅に向かうためまず不可能でしょう。
非常に残念ですが、現代世界の構図は、世界の警察官であり輸入超大国のアメリカが太陽として中心に存在し、世界中から生産物を 買いまくり、そのおかげでアメリカの周りに各国が衛星のように回っていられるだけなのです。 水星、金星はEU諸国、地球は日本、火星はロシア、木星が中国、土星がインドという感じでしょうか。
=======================================
余談
このコラムで「北京原人やジャワ原人などのホモエレクトスも出アフリカし、ネアンデルタール人も出アフリカしたということは、 ホモサピエンスが出アフリカしたのは人類の遺伝子が導く宿命ではないかとも思われる。」と書いたのですが、
何故、ヒトを含む類人猿の進化ツリー前半のギボン(テナガザル)やオランウータンは東南アジア周辺にしか棲息しておらず、 進化ツリー後半のゴリラやチンパンジーはアフリカにしか棲息していないのか?
ヒトがアフリカで発祥したのはチンパンジーとの共通の祖先がアフリカにしかいなかったからなのは極めて明白ですが、 では何故共通の祖先はアジアにはいなかったのか?何故アフリカにしかいなかったのか? この疑問に関する納得できる説明を探してみたのですが、今のところ全く見つかってはいません。
今のところの類人猿の進化ツリーでは進化の後半で、ゴリラの祖先になった類人猿はアジアからアフリカに大移動をしたことになります。 要するに、ヒトの遠い源郷はアジアだったから、人は出アフリカしてユーラシア大陸を東に進んだと言うことになります。 つまりサケやウナギが戻ってくるのと一緒で、源郷戻りが遺伝子に埋め込まれているのではないか!? では逆に、なぜ類人猿はアジアからアフリカに移動をしたのか?も依然、極めて大きな謎です。 原猿類の一部は東南アジアとマダガスカル島に生息していてもアフリカ大陸にはいないそうです。 ゴンドワナ大陸からマダガスカル島が分裂したのは6500万年前頃だそうです。原猿類を乗せたまま大陸が分裂したのですかね?! テナガザルが現れたのは2000万年前頃だそうです。オランウータンは900万年前ごろ、ゴリラは600万年前頃、 チンパンジーとホモ種が分かれたのが400万年前頃だそうです。78万年前頃に地磁気は逆転しているそうです (草創期のネアンデルタール人がホモエレクトスから進化したころになります)。 とすると900−600万年前頃に何かがあったのでしょう?こんなことに興味のある研究者はいないようです。
地質学とか気象学とかを類人猿を含む人類の進化史と照らし合わせると、もしかするとヒントがあるかもしれませんね。
とにかく解剖学的現代人類は宿命に導かれ出アフリカし、中東あたりで先輩ヒト族の地中海-中近東型ネアンデルタール人と ハイブリッドし恐らくYAP型Y-DNA「DE」と非YAP型Y-DNA「CF」に分化し、インド洋の沿岸に沿って東のアジアを目指し大移動を決行し、 古代遺伝子亜型Y-DNA「C」と「D」はアジアとオセアニアに到達しそこで棲息をしてきたわけです。 ところがY-DNA「D」の兄弟遺伝子の古代遺伝子亜型Y-DNA「E」は、せっかく出アフリカしたにも関わらずまたアフリカ (地中海南北両岸)に出戻ってしまいました。 ということはアフリカに進むことも遺伝子に組み込まれているのかもしれないですね。
では残りの古代遺伝子亜型Y-DNA「F」は、なぜインド亜大陸に留まり全現代遺伝子亜型の親遺伝子となったのでしょうか? アフリカ大陸でヒト族がチンパンジーとの共通の祖先から分化したように、インド亜大陸で現代遺伝子亜型の共通の祖先の 古代遺伝子亜型「F」から分化したのでしょう、それを実行した最も考えられる要因はアジア型ネアンデルタール人との ハイブリッドでしょう。
インド亜大陸は、パンゲア大陸でマダガスカル島と一緒だったものが、移動の結果ユーラシア大陸に接合し、 押された圧力でヒマラヤ山脈が生まれたという説明になっているようです。そのエネルギーが残っていて、 アフリカ大陸と同様の、進化や分化を後押しするパワーがあるのではないか?誰か研究してくれませんかね!!!!!。
最後に現在入手できる情報から構築できる日本人の源流ツリーです。
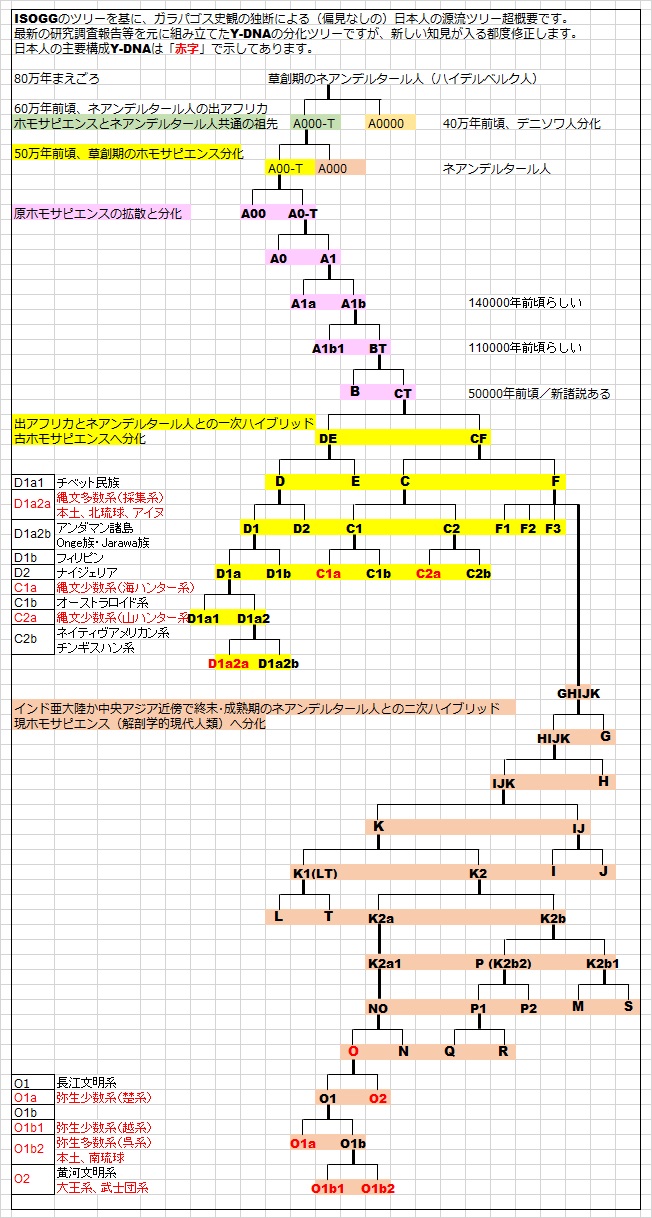
以上
表紙に戻る