インドはY-DNA「G」~「T」までの新興遺伝子全てが古代遺伝子の一つのY-DNA「F」から分化した、近代国家の故郷の亜大陸になります。 このためインド亜大陸はあらゆる新興遺伝子が分布しておりこのため何と2000以上の民族/部族/集団があると言われています。 その中でも公式に認定されている民族/部族だけでも700以上になると言うなんとも遺伝子の混沌な土地です。
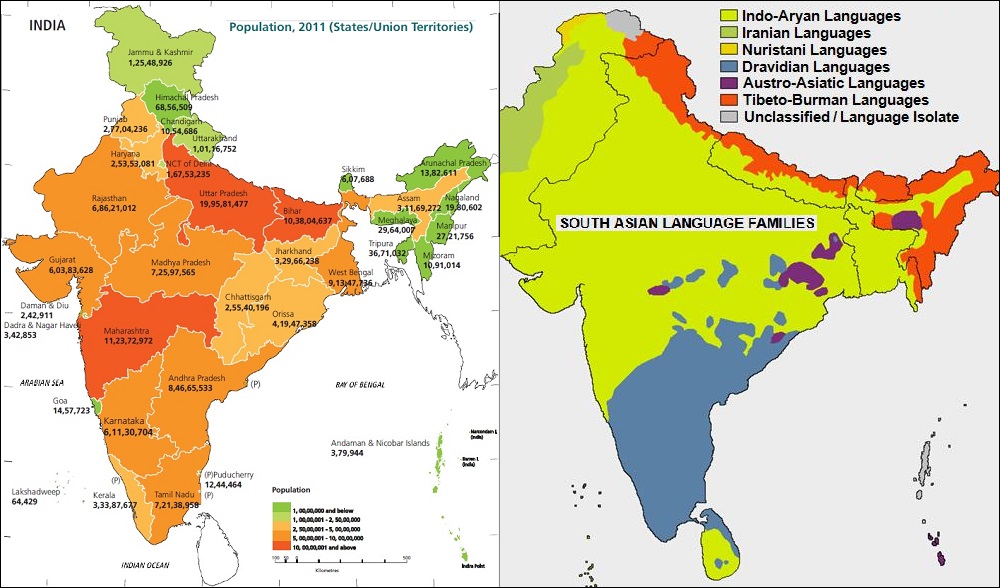
州ごとの人口データ(下左図)はありましたが、民族別の人口データは見つかりませんでした。 民族の多さもさることながら言語族と宗教に加えカーストまである3重構造になっているため、インド政府も統計には容易に出せないようです。 言語族分布図(下右図)がありましたのでご紹介します。
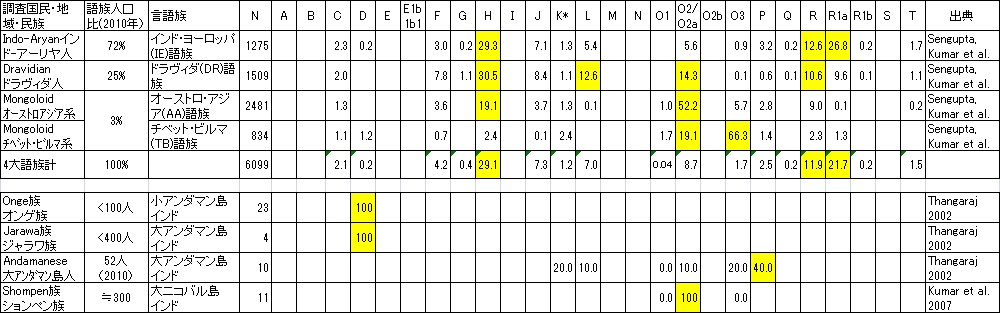
さすがにY-DNA「G」以降の全ての新興遺伝子の親遺伝子の古代遺伝子Y-DNA「F」が分化した新興遺伝子の故郷インド亜大陸です、 混沌としているのは当然なのでしょう。 と言う訳でインドの民族毎の遺伝子調査はギブアップしますが、4大言語族のY-DNA頻度データが入手できましたのでご参考にアップします。 おまけに縄文人の代表のY-DNA「D2」の祖形のY-DNA「D*」と弥生人のY-DNA「O2b」の祖形の「O2a」の情報も追記します。
カーストは部外者にはわかりにくいのですが遺伝子頻度調査で浮かび上がったのは予想通り、上流カーストは支配階級の「R1a」頻度が高く、 下層カーストは「H」頻度が高いことです。これが「H」がジプシー化した根本原因ではないかと推察出来ます。 本来なら少数民族として定着をするのでしょうが、「R1a」の支配の抑圧を嫌い定着を拒んだのかもしれません。 しかし基本的には入り混じっているので新興勢力「R1a」と先住勢力「H」の遺伝子間の交配はかなり進んでいるようです。
===============================================================
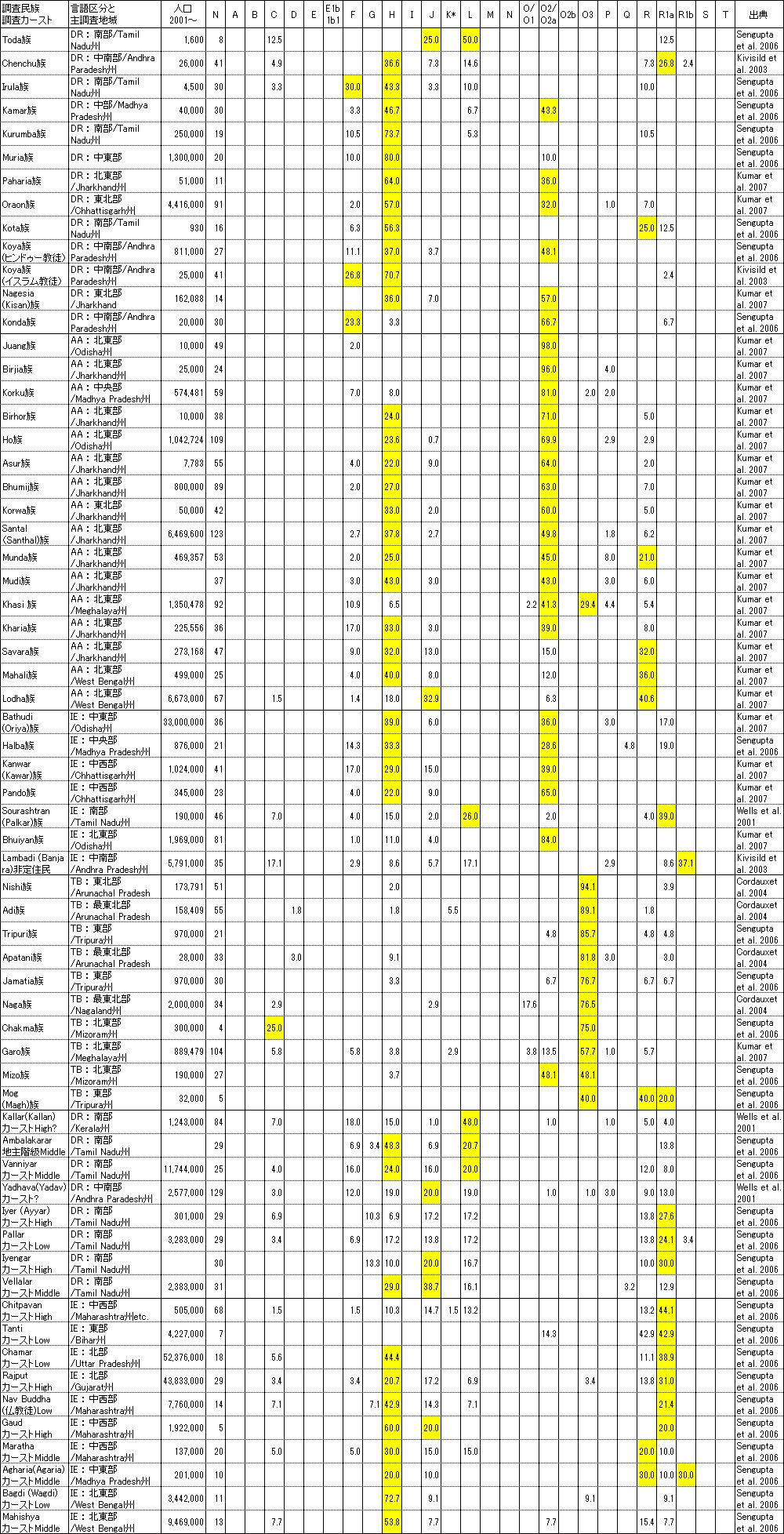
645種と書かれている指定部族(州でダブって指定されているのを除いても460種ぐらいあるとのこと)の極く一部と一部のカーストの データを集めましたのでご参考に掲載します。 残念ながら調査対象部族・カーストの人口合計も2億人強に過ぎず12億人と言われるインド人口の17%程度に過ぎません。
あまりに一部のデータなので、これがインドだ、と言える物はないのですが、ドラヴィダ系、インドアーリアン系、 オーストロアジア系とチベット・ビルマ系の一般部族の特徴は良くわかると思います。 日本人の弥生系の主役Y-DNA「O2b」/「O2b1」の兄遺伝子のY-DNA「O2a」水田稲作農耕民がオーストロアジア系民族のみならず ドラヴィダ系やインドアーリアン系にも深く入り込み水田稲作農耕をインド亜大陸に広く広めたことがよく理解できます。
ところが一般部族とは異なり、カースト集団になると状況は一変し、下層と差別されてきた極東モンゴロイド遺伝子Y-DNA「O2」も「O3」も カースト集団からは誤差程度にしか検出されません。Y-DNA「R1a」を基盤とするインドカースト集団がいかにモンゴロイドを下層民として 差別し集団から排斥してきたか過去の歴史が結果としてこれほど見事に現れるとは思いませんでした。日本人は世界3位のGDPでインド人と 付き合えていますが、中国が人口力でGDPを上げた様に、人口で中国を近い将来追い越すと確実視されているインドがGDPで日本を 凌ぐようになると、途端に今の中国のように、日本いじめを始める可能性もあるかもしれません。
それにしても部族名がありながらも遺伝子が純系の部族はもはやなく、インド亜大陸の長い歴史の間にY-DNAは相当混じり合い、 さすがに全ての近代Y-DNA亜型が発祥した亜大陸の奥の深さが感じられます、あまりにも深すぎてこれ以上追うのはむりなので止めますが、 興味のある方は更に深堀りしてみてください。
以上
表紙に戻る
============================================================
日本人の遺伝子を構成するY-DNAもmtDNAも全てがこのインド亜大陸で分化しサハラ以南のアフリカ以外の全世界に拡散をしたと 言われています。
つまりホモサピエンスの故郷はアフリカ大陸東部ですが、現代人の故郷はインド亜大陸なのです。
遺伝子調査を進めると、民族は基本的に言語族と対応していますが、過去の歴史でから遺伝子の移動拡散もかなり追えるようです。
1. インド亜大陸で最も古い遺伝子は全新興遺伝子の親の古代遺伝子Y-DNA「F」です。古代遺伝子「C」と「F」は 出アフリカ後の中東地域でネアンデルタール人と交配しネアンデルタール人が数十万年かけて獲得したエピジェネティクス形質を 一気に手に入れたようです。 赤毛碧眼・金髪・彫深顔・色白肌・大柄等々は全てネアンデルタール人のオリジナル形質と思われます。
2 「F」の長男遺伝子のY-DNA「G」はコーカサス遺伝子と呼ばれるようにインド亜大陸には留まらずコーカサス一体に移動をしたようです。 コーカサスの高原・山脈地帯が好みだったようです。
3. 二男遺伝子の「H」はインドの重要な遺伝子として今も残っていますが、その後のインダス文明人やインド・アーリア人の興隆で インド亜大陸の主役ではなくなりました。一部は定住できずに放浪するジプシー民族となりました。
4. 次の「I」はどんどん西に進みヨーロッパ大陸でクロマニヨン人として遺跡を残し、氷河の後退とともに地中海沿岸から北上し後に ノルマン人として活躍することになりました。一部はヴァイキングとなりヨーロッパ大陸沿岸を荒らしまわり、一部は現ロシア地域に 進出し公国等を興し、子孫にトルストイを残しました。
5. 次の「J」はセム族遺伝子としてまたメソポタミア文明遺伝子として「I」同様西に進みヨーロッパ大陸に最先端の農耕文化をもたらし、 その後アラブの主要遺伝子となり地中海両岸の主要民族となっています。
6. 次の「K」は移行段階の遺伝子でインド亜大陸に留まったようですが、「L」以降の全ての遺伝子の親遺伝子となりました。
7. 「L」はインダス文明の遺伝子と認知されたようです。と共に「L」を主遺伝子とするドラヴィダ人がインダス文明人の子孫 であることも認知されつつあるようです。
8. 「T」は遺伝子頻度が低く本来「L2」でも良かった子亜型タイプなのですが、米国第三代ジェファーソン大統領の遺伝子であることがわかり、 主要ハプロタイプに格上げされました。ところが「S」まで既に埋まっていたため「T」となったのです。
9. 「M」はインド亜大陸を東に進みスンダランドから更にサフールランドに進み、ニューギニア・メラネシアの主要遺伝子になりました。
10. 「N」は移行タイプの「NO」時代に東進し華南に移動しそこで「O」と分化し、更に北上しシベリアに住み付きシベリア系遺伝子 となり寒冷地適応しましたが、多くが北ユーラシアを西進しバルト海・スカンジナヴィア周辺に住みつきウラル系遺伝子ともなりました。
11. 「O」は言わずと知れた極東遺伝子です。先進だった「O1」(楚系?!),「O2a」(越系?!),「O2b」(呉系?!)が長江文明を興し、 後進だった「O3」は北上し自然が厳しい黄河流域で黄河文明を興しその後中華大陸の覇者になり、現在は極東のほとんどの先進国の 支配階級遺伝子として君臨しています。現在は世界の覇者「R1b」に覇者交代を目論み戦いを挑んでいます。
12. 「P」は基本的にはインド亜大陸に留まりました。やはり移行段階の遺伝子で「Q」と「R」の親遺伝子となりました。
13. 「Q」は大冒険者遺伝子で北上しベーリング陸橋を渡り南アメリカ大陸の南端まで拡散し、南北アメリカ大陸の長い間主役の 遺伝子でした。
14. 「R」はインド亜大陸に留まりましたが、その中から「R1」が分化し更に「R1a」と「R1b」が分化し、「R1a」はインド・アーリア人 としてインド亜大陸の主役となり、一方北西に「R1b」と共に移動した集団はスラブ人としてロシアに留まり主役になりました。 しかし更に西進した「R1b」の中で純血を好んだ一部はケルト系民族としてヨーロッパの先住民となり、 多くは「I」や「N」と交配しケルト・ゲルマン系となり、古代遺伝子「E1b1b」と交配した集団はローマ・ラテン系としてローマ帝国を 興隆させました。いずれにせよ現代ヨーロッパの主役であり、「R1b」は世界の覇者の遺伝子です。
15. 「S」はハプロタイプとしては不思議な存在です。何故「S」が分化する必然性があったのか全く謎ですが、しかも何故 ニューギニアに存在するのか?ニューギニア・オーストラリアのサフール大陸には古代遺伝子「C2」が少なくとも4万年前には マグロ漁を行っていたことが報告されています。そこに新興遺伝子の「M」がインド亜大陸から移動をしニューギニアから メラネシアに進出しました。過酷な土地なのでそれ以上の遺伝子は必要なかったはずなのに突如「S」が出現したのです。 しかも「C2」や「M」同様ニューギニアとその東の太平洋の島々のみに生存しているのです。今のところの仮説はネアンデルタール人の アジア型のデニソワ人がシベリアから南下する途中で「R」と交配し分化したのではないかと言うものです。
インド亜大陸の遺伝子調査で判明したことの中で、日本人にとってもっとも重要なことは;
1. インド亜大陸そのものではなくインド洋上のアンダマン諸島には縄文人Y-DNA「D2」の祖形「D」の直系子孫 Y-DNA「D*」100%のJarawa族とOnge族が残っていること。
2. アンダマン諸島の南に続くニコバル諸島には長江文明の子孫で弥生人呉系Y-DNA「O2b」の祖形「O2」の直系子孫で 越系Y-DNA「O2a」100%のShompen族が残っていること。
3. 大野教授が日本語のルーツと発表し話題になったドラヴィダ系タミール人はなんと平均14%もの「O2a」頻度があることや、 他のドラヴィダ人には「O2a」98%もの純系にちかい部族もいること。 この長江文明越系子孫の「O2a」遺伝子集団がタミール語を含むドラヴィダ語族に水田稲作農耕の語彙を持ち込み、 日本語の稲作関連語彙と強い相関を持つのが当然だったのです。 黄河文明との戦いに敗れた長江文明の子孫はそのまま留まり黄河文明に飲み込まれた集団もあれば、 ある越系「O2a」集団は南へ逃げ遠くインド南部まで逃げ、先進の水田稲作農耕という食糧生産法を持つために インド亜大陸でも現在まで生き延びることに成功したのです。一方呉系「O2b」集団は北へ逃げ満州に遺伝子を残し 稲作適地を求め南下し朝鮮半島南部に30%もの遺伝子頻度を残し、更に南下した一部は日本列島に入り弥生文化を残し 30%強の頻度を現在でも残したのです。
以上
表紙に戻る