表紙に戻る
本ブログは日本の御用学者や専門家の恣意的な報告に惑わされずに、グローバルに発表されインターネットで入手できる 論文の成果から客観的に日本民族のガラパゴス化の起源を探るのが目的です。 また素人にありがちな近視眼的な狭い視野に陥るのを出来るだけ防ぐため、学際的な本を読み大所高所からの判断ができるように 自己啓発をするサイトでもあります。
日本民族に関連する遺伝子頻度のデータを発表しているほとんどの原著論文はPDFで手に入りましたが、1誌だけ有料でないと 閲覧出来ない学術誌がありました。残念ですね。
それはさておき、入手できる限りの引用される論文から集めたY-DNA頻度リストを掲載しましたので興味のある方は眺めてください。
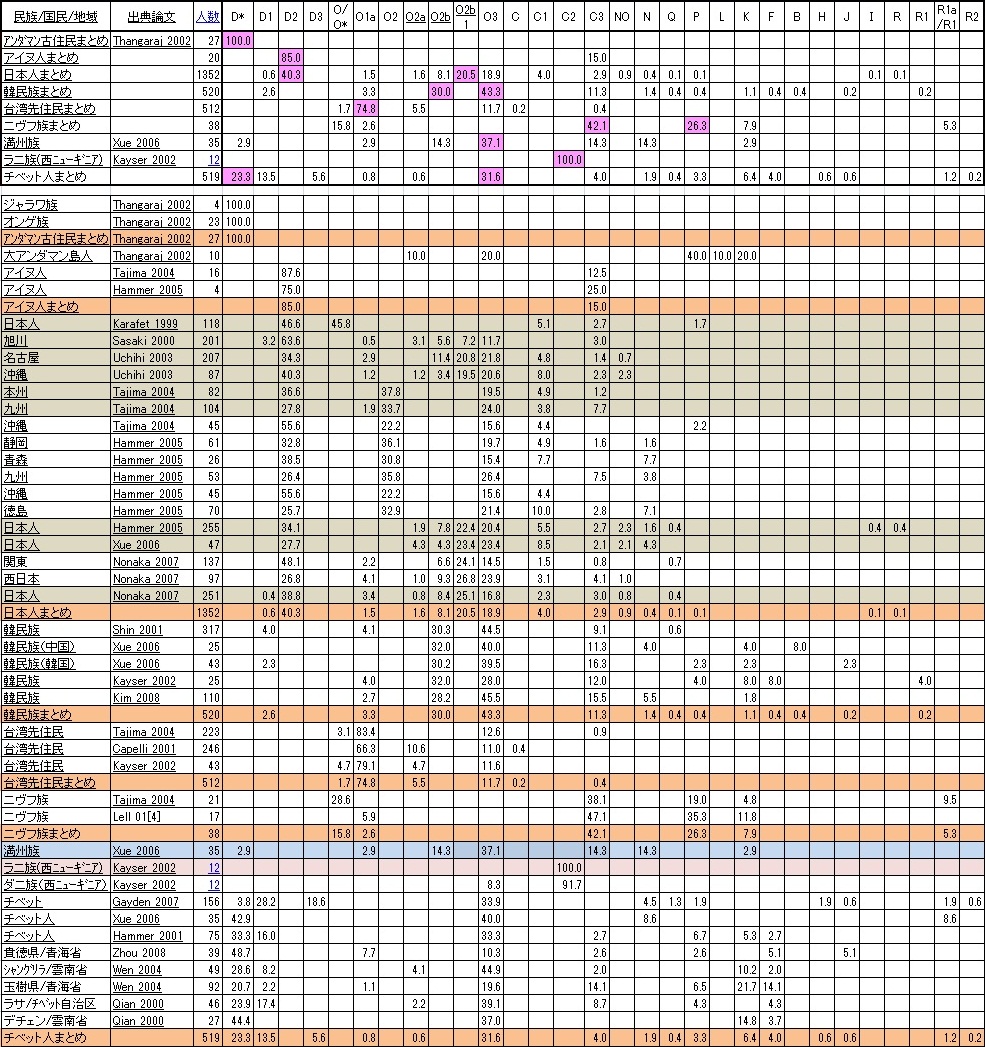
1. 縄文主力系Y-DNA「D2」のオリジン、Y-DNA「D*」のアンダマン諸島/オンゲ族、ジャラワ族
後世にインドからの統治者の来島やカレン族の島流しなどで遺伝子は荒らされた、と思っていたのですがThangarajの論文で Onge、Jarawaの2部族はY-DNA「D*」を100%維持し、 出アフリカした現代人類の世界最古の姿をとどめている純系の集団であることがわかりました。縄文人の主力集団のY-DNA「D2」の祖形になり、 日本人に最も多いジャガイモ顔でパッチリ二重瞼ののルーツになります。女性は典型的な安産型の大きな臀部でいわゆるQPちゃん体系です。 ホッテントット族が典型的ですが臀部に脂肪としてエネルギーを蓄えるのです。出アフリカ時の飢餓状態だったアフリカ・サハラ時代の 名残をとどめているのです。 そのくらい古代の遺伝子なのです。
しかし50000年もの間純系を保てたのは定住先の小さなアンダマン諸島で外来者を殲滅する部族の習慣だったのですから 何とも複雑な感じがします。 もし縄文人主流の「D2」が純系を保つために少数派の「C1」や「C3」との交配を拒んでいたら今の日本人は存在しなかったかもしれません。 スンダランドか東シナ海-黄海ランドが広大な土地だったため、棲み分けが可能だったからではないかと思います。
歴史を見ると男子の侵入者はせん滅しますが、アマゾン等の他の先住民と同様近親婚による遺伝的障害から部族消滅を防ぐため 女子は他の近隣部族から新しい遺伝子を導入してきたようです。 こうして部族を維持してきたのです。
2. 縄文系海洋性ハンターY-DNA「C1a」の兄弟Y-DNA、Y-DNA「C2a」の西ニューギニア/ラニ族
ラニ族はY-DNA「C2」を100%維持しています。縄文人の一員の海洋性ハンターY-DNA「C1」に最も近い弟遺伝子になります。 「D2」とは異なるもう一つの出アフリカした世界最古の現代人類遺伝子である「C1」の縄文時代の姿が彷彿とされます。 「D2」のジャガイモ顔集団に対し目が窪んでいる奥二重目のいかついソース顔集団のルーツで、「D」とは全く異なる風貌です。 Y-DNA「CF」からY-DNA「D」,「E」以外の全ハプロタイプが発生しています。中東でネアンデルタール人と亜種間交配をし分化した Y-DNA「DE」と「CF」ですが、 「CF」から分化した「C」はネアンデルタール人の形質を受け継ぎ、その結果黒色は褐色に薄まり、風貌もネアンデルタールがかった顔に 変化したものと思われます。 似た人は周りに以外と見かけるので、日本人にすっかり定着しているようです。
3. 縄文系Y-DNA「D2」の兄貴分Y-DNA「D1」のチベット民族
チベット人はY-DNA「D1」と「D3」を残す縄文系日本人の兄弟遺伝子集団ですが、「O2b」や「C1」が全く存在しないことや、 大陸の中にいるため「K」「F」など 日本人にはない遺伝子タイプが存在することが大陸に住むチベット民族の特徴になっています。早くから「O3」など周辺の漢民族や 異遺伝子集団との交配が頻繁に行われてきたため、 代表的なチベット人はどんな特徴かは全くわかりません。しかし民俗学の本にはチベット中心域には小柄な人が多いそうなので Y-DNA「D1」の持っていたネグリート的形質を 濃く残している集団がかたまって居住しているようです。
チベットはチベット密教の国で、長野の善光寺は密教系のお寺です。どうやらY-DNA「D」は密教的な神秘的な神事に魅かれる 遺伝子なのかもしれません。 アンダマン諸島では、まだ宗教が確立する段階になってはいないようです。
4. 韓民族
北朝鮮のデータは当然ありませんが、中国の朝鮮族のデータで推測することが出来ます。日本と近くて遠い存在であることが はっきりとわかります。 似ている部分は長江文明の流れ汲む呉系の水田稲作農耕民遺伝子の「O2b」を共に30%近く持つことです。これが韓民族に感じる親近感の原因です。 遺伝子の約1/3が共通すると言うことは極めて大きいと思います。一方似ているようで違うのが「O3」と「C3」の頻度が日本より高いことです。 特に朝鮮半島史上1000回にも上るという中国王朝の侵攻・侵略・支配のせいで朝鮮半島の遺伝子基盤は、 本来の北部の「C3c」や南部の「O2b」から「O3」に変化しています。 「D2」が遺伝子基盤の日本民族との決定的な違いになっています。
日本人が一般的に感じる一重瞼の韓国人の姿は寒冷地適応・黄砂適応した黄河系「O3」とツングース系「C3c」の特徴です。 しかし釜山や済州島など南になるほど二重瞼が増える理由は「O2b」が南に分布しているからに他なりません。30年以上前に仕事で釜山に入り、 済州島も訪れそのままバスで各地のユーザーを訪問商談しながらソウルに入りましたが、北に向かうほど、 一重瞼一色に変化してゆくことをはっきり覚えていますが、 遺伝子調査は朝鮮半島人の出自の違い、最大の黄河系漢族系、二番目の長江系呉系、三番目がツングース系であることを 見事に解明してくれました。
Y-DNAハプロタイプがもともと異なるのです。北部ほど黄河系漢族系「O3」が多いのは楽浪郡など中国の古代帝国の組織が置かれ、 長く中国王朝の一部として占拠されていたところだからです。南方は水田稲作地帯で長江文明の子孫(恐らく呉系民の)が 住み着いた場所だったのでしょう。 だから韓半島極南部は日本の原風景とかなり似ている感じがするのでしょう。
5. 台湾先住民
台湾の一般市民は漢民族の「O3」系ですが、先住民は長江文明・楚系の「O1」75%の遺伝子集団なのです。日本列島とは相当異なるのです。 にもかかわらず日本人が台湾人に感じる親近感はルーツの福建省など華南出身の台湾漢民族が持つ黄河系の中でも 特に長江文明との交配系の漢民族「O3」遺伝子に起因するのでしょう。
日本人の中に「O1」は全体では少ないのですが、中国地方のようにY-DNA「O1」が突出して多い特殊な地域があり 日本人には違和感がない遺伝子であることも親近感の一因でしょう。
中国地方は「O2b」「O2b1」も多く、「O1」と合わせなんと50%も占める日本列島の中で独特の長江文明系が主流の土地柄 (水田稲作農耕が発達していたはずで恐らく「瑞穂の国」の原型の土地でしょう)なのです。 古代に独特の勢力を誇った吉備氏は恐らくそのY-DNA「O1」と「O2」の長江文明系大集団だった可能性が極めて高いのです。
スンダランドから北上した「D2」「C1」は当時陸続きの台湾部分には住み着かずにそのまま日本列島部分に進んだようです。 理由は勿論わかりませんが、 もっと北を目指していたからかもしれません。
6. ニヴフ族
Y-DNA「C3c」純系集団は既に存在しませんが、最も頻度が高いのはオロチョン族61%~91%です。しかし寒冷地適応をして フラットフェース化してしまっているため、 ナウマン象を追って日本列島に南下してきた頃の寒冷地適応する前の「C3a」本来の姿は全く推測もできません。
現在のオロチョン族の外観はその辺にいる日本人と同じです。オリジナルの「C3c」は西ニューギニアの「C2a」ラニ族のような 彫の深いソース顔だったはずですが、 寒冷地適応の結果、遺伝子の発現が変化しフラットな顔に変ってしまったのです。 (「O3」は寒冷地適応に加え黄砂適応も加わっているそうです。)
一方北海道に侵攻し「D2」を征服しオホーツク文化を立ち上げたのは古代ニヴフ族になります。そして原アイヌ「D2」と交配し 現代アイヌ民族を作り上げました。 熊祭りや民族衣装など主だった風俗習慣は残念ながらほとんど支配階級だった古代ニヴフ族のものです。寒い土地になってしまったため 南から北上した「D2」本来の風俗習慣は 何も残っていないのではないかと思われます。それはアイヌ自身ももはや区別できないでしょう。ユーラシア大陸に残っている 現代ニヴフ族にはない風俗習慣がもしあれば 「D2」オリジナルな可能性はありますが...
7. 満洲族
満洲族はツングース族の有力な一員(女真族)ですが、何となく親近感があるのは「清」の王族であることと、大日本帝国が デッチあげた満洲国の構成部族だったこと、 愛新覚羅の名前が持つ悲劇が話題になった近代の状況がありますが、水田稲作農耕遺伝子「O2a」が弥生人の祖先である「O2b」に 分化した土地であるため、 今でも「O2b」が約15%含まれることも大きい要因でしょう。遺伝子的に15%も共通なのです。 但し極東遺伝子度調査でも触れましたが、この満州族に見られるY-DNA「O2b」のオリジンは、1744年編纂の「八旗満州氏族通譜」で 満州族中に43姓の朝鮮族姓が既に記載されているほど 満州族中に深く浸透している朝鮮族の 伝えてきたY-DNA「O2b」である可能性が極めて高く、満州族がもともと持っていた 遺伝子ではない可能性が大です。 恐らく朝鮮の原点と考えられている箕子朝鮮や衛氏朝鮮の子孫が後世満州族に取り込まれたからかもしれません。
8. アイヌ民族
アイヌ人がアンダマン古住民に次いで「D」が極めて高頻度の集団です。黥面など顔面装飾文化を維持してきたことがアンダマンと 共通の古民族であることを示していましたが、 前にも説明したように日本本土のY-DNA「D2」は「C1」と共に日本列島を北上し青森で留まり後に縄文文化を花咲かせた集団ですが、 アイヌの「D2」は日本列島には入らずに「D1」とともに韓半島からユーラシア大陸に渡り、「D1」が中原の最初の民族になったのに対し、 日本海沿岸を北上しサハリンやカムチャッカまで到達し、 一部が南下し陸続きだった宗谷海峡を渡り北海道に分布したと推測されます。元は同じ集団ですが、途中の経路・経緯が異なり、 「C1」が存在しなかったこと、寒さが厳しかったこと、等々.. で本土とは全く異なる歩みになったのでしょう。「D2」縄文人とオリジンは全く同じ遺伝子でも、その後の交配環境が異なると これだけ違ってしまう典型的な見本です。
9. 日本民族
日本人は世界でも指折りの多種類の遺伝子タイプを持つ集団ですが、果たして「日本民族」と呼べるような典型的な集団に なっているのでしょうか? 結果として狭い島国であることが幸いし「O3」以外の外来遺伝子の撹乱が全く起きなかったこと、「O3」は大和朝廷族や武士族として お上の支配に徹してくれたため、 縄文-弥生の先住混成集団は地味ながらも列島各地で風俗習慣を守ることができたようです。
各研究者の結果では北海道と沖縄(北琉球)が「D2」頻度が高いのです。北海道は屯田兵が中心で開拓がなされ「D2」系の 貧農農民が多かったせいでしょう。 「D2」は寒さに対し我慢強いのでしょう。沖縄に多い理由は先に触れましたが、「D2」「C1」混成部隊は、琉球列島を通らずに 当時の列島部分の北側の海の北岸を進み一部は日本列島部分を北上し、 一部は南下して琉球列島部分に入り、南琉球との間の当時の海峡に阻まれ北琉球に留まったためそこに住み着いた可能性が大きいのですが、 (ただし「C1」は海洋性だったため舟で北上し琉球列島に入り港川人の遺跡を残した可能性もあります)。 そのため「O2b]が極めて多い南琉球ではもともと「D2」が住んでおらず、後世に尚王朝から移住した「D2」の痕跡程度の頻度しか ないのでしょう。薩摩藩時代の混入は考えにくいです。
「O3」は九州に多いのです。朝鮮半島を追われた負け組の武装侵攻集団の一員つである大和朝廷族が書いた日本書紀が 日向から始まるのはハプロタイプ頻度からみて当然のことだと思われます。 大和朝廷族が攻略し支配していた奈良や京都の遺伝子頻度を誰か調べてほしいですね(実はY-DNA調査の空白地帯なのです)。 断トツの「O3」頻度になる可能性があります。同じくY-DNA調査空白地帯の大阪の独特の文化も縄文-弥生系とは全く思えないので 「O3」や「C3c」の可能性が高いのです。東京はゴチャ混ぜです。
奈良、京都、大阪の遺伝子を調べると日本民族の「O3」頻度は飛躍的に上がる可能性が実はあります、 「C3c」も出てくる可能性があります。もしそうならないとすると、 少数の部隊で日本列島を攻略してきたことになります。勿論武装侵攻集団なので狩りのための武器しか持たない狩猟採集・農耕混成集団の 縄文-弥生人を蹴散らし制圧しながら東進したのでしょう。
ただし、既に上陸していた武装侵攻「O3」部隊は他にもあり、物部系の饒速日尊(にぎはやひのみこと)集団もその一つだったでしょう、 最終的に勝ち残ったのが大和朝廷族に成ったのでしょう。
10. 「C3」の違い
日本列島は古代亜型Y-DNA「C」にとって特別の土地です。
最初の子亜型「C1a」が維持されているのが日本のみ。
「C3」の最初の子亜型「C3a」も日本のみ。
古代遺伝子「CF」から分化した古代遺伝子亜型Y-DNA「C」,「F」は日本人にとってどんな存在だったのか、 実は「O」亜型は「F」の子孫なのです。日本人のY-DNA遺伝子の約40%はYAP系のY-DNA「DE」から分化した「D」ですが、 その他60%は全て「C」と「F」つまり非YAP系「CF」の子孫なのです。つまり「CF」系の方が頻度が高いのです。 現代人類のY-DNA遺伝子の最初のハプロタイプつまり 古代シーラカンス遺伝子「C」,「D」,「E」,「F」のうち「C」,「F」と「D」で日本人は構成されているのです。
欧米人の研究によると現代人の遺伝子に取り込まれたネアンデルタール人遺伝子は1-4%を占めるそうです。 「D」,「E」は交配しても出アフリカ時の現代人類の オリジナルの形質、色黒/小柄等、を保っているのに対し、「CF」は形質をかなり受け継いだらしいのです。 日本人の持つ「CF」系遺伝子が60%を占めると言うことは、 日本人にもネアンデルタール人の遺伝子が受け継がれている、と言うことに他なりません。
では、日本人特有の「C3a」以外は「C」の子亜型は?
ニヴフ族は「C3c」 満洲族は「C3d」 チベット族は「C3e」
では「C3b」はどこへ行ったのでしょうか? 陸続きだったベーリング海峡を渡りアメリカ大陸に上陸しました。 ナデネ語族、アルゴンキン語族などのネイティブアメリカン (アメリカインディアン)をY-DNA「Q」と共に構成しています。残念ながら純系の「C3b」は存在しません。
11. 縄文土器
縄文土器は千島列島やサハリンなど旧アイヌ民族地帯からまだ発見されていません、しかし中米に縄文土器によく似た土器が 出土し遺伝子構成は「Q」と「C3b」です。 「C1」は貝文土器の作成者と考えられていること、土器を使って煮炊きをするのは穀物より大型動物の肉の方が要求度が高い... ことなどから推測すると縄文土器はハンターであった 「C1」「C3a」が主役となり作成し、人口比が大きかった「D2」は使用者だったような気がします。 勿論自分たちでも作成するようになっていったのは当然でしょうが...。
12. ガラパゴス性
となると「創意」発案能力が高いのはY-DNA「C」系のような気がします。 「工夫」はもっぱら使用する側の「D2」が得意だったら面白いですね。 しかし、ガラパゴスの真髄は前にも書きましたが、大和朝廷族が中国から連れてきた「O3」官僚集団と技巧集団だと思います、 遠い異国に来て本家の中国の高度な文化に負けない文物を作り出す間に 徹底的に知識。技術を極める集団になって行ったはずです。
つまりガラパゴス的民族性の起源は、文化の本家「中国」を追い越そうと突き詰めてきた結果なのだと思います。 そこに「C」の「創意工夫」が加わり、 更に時代が下がって江戸時代の鎖国中にとことん突き詰める民族性が昇華熟成され強固なガラパゴス性が形成されてきたのではないでしょうか。 そして明治時代に入り、 かって本家中国に追いつけ追い越せと頑張った奈良天平飛鳥時代以来の、西欧文化に追いつけ追い越せと頑張った結果が、 日本民族のガラパゴス性の直近の起源です。 1000年以上の気合の入った筋金入りです。Y-DNA「R1b」なんかに負けてたまるか...ですね。
識者・評論家はすぐグロ―バル化と言いますが、西欧文明の原動力のY-DNA「R」遺伝子が最近の外国人との国際結婚以外、 全く混じっていない日本人が、「R」のまねをするのは大変です。 「D」「C」「O」混合集団の独特のやり方があってもおかしくはないはずです。今や世界最大の人口を持つ遺伝子になった「O」です。 アジアのスタンダードをグローバルにできる可能性はあります。 そのためにはかっての本家中国と組む必要があるかもしれませんが...ですね!?
以上
表紙に戻る