地方別集計
県別データ
以上
表紙に戻る
●全国データまとめ
日本列島のY-DNA出現頻度とmtDNAの出現頻度は既に報告していますが、ここでは同じGlobal論文からのデータを使っていますが、 県別や地方別のデータから積み上げ、 県別の人口比や地方別の人口比で按分加算したデータとなっており、単純加算した4-5.より統計上はより良い試算データとなっています。 XueとTajimaの論文は子亜型の分析がまだなのですが亜型単位では充分なのでデータに加えました。 先駆的な論文として必ず引用されるHammerも共著のKarafet et al.1999のデータは残念ながら「O」が「O2」と「O3」に分離されておらず、 日本人のデータとしては極めて不十分なので試算データには含めませんでした。
いずれにせよ、日本人が縄文文化系、呉系弥生文化系及び征服者プロト漢族系の3大遺伝子起源から成り立つことは 完全に間違いないようです。
縄文系はさらに「D2」、「C1a」及び「C3a」の3亜型から構成されることも間違いないようです。当ガラパゴス史観では この3タイプは日本列島で1万年間交配しあい、 それぞれ子亜型に分化していますが、たため、この日本列島独自のY-DNA亜型が、朝鮮半島には欠落しているため遺伝子が 日韓で大きく異なることがわかりました。
Y-DNa「C1a」は現在ニューギニアやオーストラリアでオーストラロイドと称される集団の中の海洋性ハンターで、 各地に残る鯨漁などの基地は「C1a」の居住地の名残でしょう。 Y-DNA「C3a」もナウマン象など内陸型大型獣ハンターで現在のアメリカインディアン「C3b」(ネイティヴアメリカン)が 最も近い遺伝子で九州を中心に日本列島に拡大しています。 山岳系集団の起源のはずです。このY-DNA「C」はネアンデルタール度が「D2」より高く、 日本列島の彫深・鼻高・面長顔で二重瞼か奥二重のソース顔イケメンの起源遺伝子のはずです。 (「D2」は二重瞼なのですがゴロッとしたジャガイモ的丸顔に近い顔つきです)
遅れて日本列島にボートピープルとして耕作適地を求めて長江→満州→朝鮮半島から南下した呉系長江人の弥生系のコア遺伝子Y-DNA「O2b」は日本列島に上陸後「D2」と密に交配し、 稲作が意外に早く東北まで展開された要因となり満州と朝鮮半島には存在しない「O2b1」に分化し、「D2」もさらに新しい子亜型に分化しています。
このように日本列島独自のY-DNA遺伝子タイプが日本列島の大半を占めることが、 似ているようで実はかなり異なる日本人と韓国人の違いとなっています。 しかし「O2b1」に分化していないオリジナルの呉系長江人「O2b」も日本列島には或る程度存在することが 韓半島との過去の同一呉系水田稲作農耕文化圏に基づく「近さ」の要因になっています。 Y-DNA「O2b」と「O2b1」も水田稲作農耕民の一部として一緒に日本列島に渡ってきた楚系Y-DNA「O1a」や越系「O2a」も 全て二重瞼の面長顔でも標準的な彫浅のしょうゆ顔日本人です。
そして日本列島で3番目を占めるのが大和朝廷族や武士団族などの朝鮮半島の中華王朝出先機関から生き残りに負けて 日本列島に流れてきたY-DNA「O3」プロト漢族の武装侵攻遺伝子群です。 この「O3」は黄砂適応・寒冷地適応をしていたらしく面長顔の彫浅の一重瞼で極めつけのしょうゆ顔でしょう。 一緒に日本列島に渡っていたと考えられるY-DNA「NO」や「N」、「P」,「Q」等も 寒冷地適応組なので彫浅一重瞼組と考えられます。
Y-DNA「O3」は黄河文化直系のプロト漢族、「O3a1」は華北の周辺民族との交配で生まれた黄河系(華北系)漢族、「O3a2」は 黄河系漢族と長江文化華南系との交配漢族になります。 漢族は単独遺伝子ではなく、約50%強の「O3XX」子亜型群をコアに現在少数民族となっている「O1」「O2」「F」「P」「NO」N」「C3c」など 50以上の民族との交配の結果生まれた 複雑な遺伝子構成の集団ですが、支配階級は「O3XX」なので「O3」で代表されます(面白いことに他の遺伝子との交配を嫌った 純粋「O3」の少数民族もいるのです)。
いずれにせよ日本列島で検出できる「O3」の子亜型は全て中国でも検出できます。そして朝鮮半島の「O3」とも全く同じです。 つまり「O3」漢族は支配階級として君臨し 日本列島先住民の縄文-弥生集団との交配を行ってこなかったことを意味します。自分たちはエスタブリッシュメントとして 先住縄文-弥生集団を差別していたようです。
この「O3」は朝鮮半島の支配階級でもあるため、朝鮮半島も日本列島も漢族系の支配した土地であったことがよくわかります。 日本列島は残念ながら今でも「O3」封建体制のままと言っても過言ではないでしょう。 それでも基本的におとなしく、黙々と働き、年長者を敬い、和をもって貴しとなす縄文-弥生遺伝子集団は、 「O3」には支配される傾向にあるのでしょう。
以上
表紙に戻る
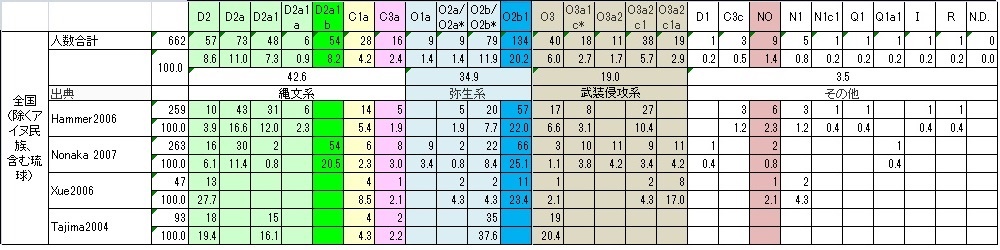
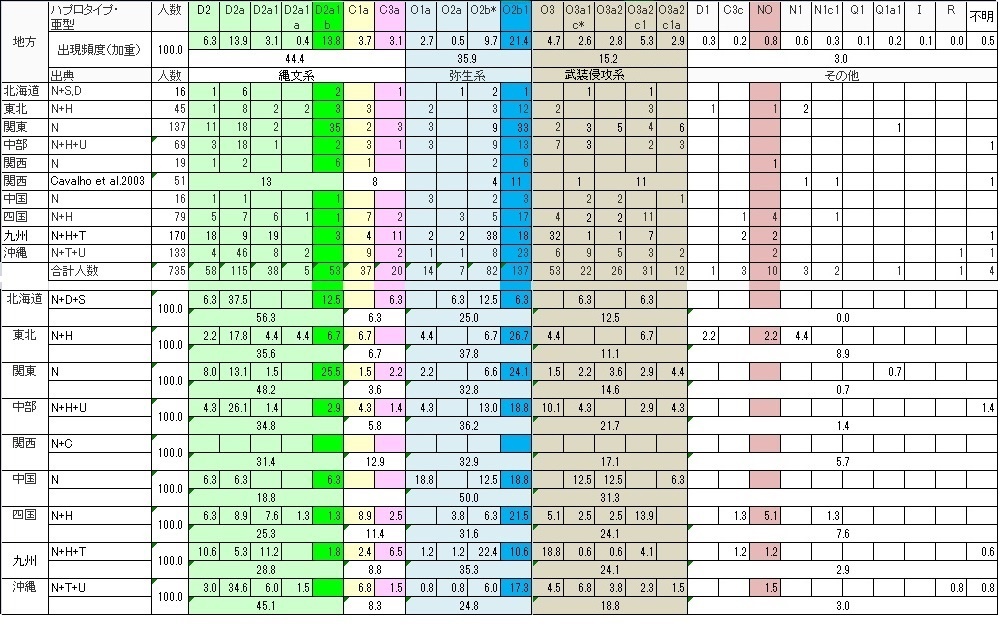
●地方別データまとめ
県別のデータではサンプル数が少なすぎ県民論は追えませんでしたが、地方別になると、地方単位で調査したGlobal論文もあり、 それなりに追える場面も出てきました。 それでも関西圏などまだサンプル数が少なく、調査結果に偏りがあり過ぎ論じるのは難しいようです。
Webで関西のGlobalデータを見つけました。このCavalhoデータは亜型の設定がやや大雑把のため他のGlobalデータと 比較をしにくいのですが、逆に縄文系、弥生系、侵攻系の 大くくりには使えるため加えました。特徴はY-DNA「C」が高めなのです。関西(近畿)圏は和歌山の太地の鯨漁などで予想される 海洋性ハンターY-DNA「C1」の居住地、 まだ修験道などで予想される内陸型ハンターY-DNA「C3a」の居住地でもあるはずです。Y-DNA「C」が多めなのは充分納得できます。 またNonakaデータではなかった「O3」もあり、むしろCavalhoデータの方が納得できるくらいです。
県民性ではまだ論じることはできませんでしたが、地方性では傾向が見えてきました。
北海道は、明治維新後の屯田兵など開拓民に縄文系が多かったことが言えます。貧民が多かった縄文系は海外移民と同じ発想で 新天地として北海道に開拓民として入ったことが予想されます。 当然支配階級の「O3」は厳しい自然の土地には政府の役人以外は行かなかったのではないかと予想できます。
東北も、縄文-弥生の土地ですが、寒冷地適応系と考えられるY-DNA「NO」、「N1」「N1c1」の比率が最も高く、 寒さに強かったため適応できたのでしょう。 東北アジアに住んでいた時から高緯度地適応で色白だったはずなので、東北に多い色白美人の要因かもしれません。 これまで蝦夷はY-DNA「C1a」ではないかと推測していたのですが、 「NO」N1」「N1c1」の東北アジア起源集団も蝦夷の一員であった可能性が見えてきました。 この東北アジア集団は韓半島を追い出された「O3」と一緒に対馬海峡を渡ったはずですが、 故地に似た気候を求めて日本列島を北上した可能性があります。「O3」は全地方で最低でした。 奈良時代、平安時代と支配階級の「O3」は蝦夷との抗争がありましたが、 積極的に住むことはしなかったようです。
関東は意外に縄文系Y-DNA「D2」頻度が高い土地でした。新開拓地の北海道を除くと、沖縄以上の「D2」地方になります。 しかも人口比率では日本列島最大なので、 縄文系の頻度を押し上げることになります。海や山が充分ではないため縄文ハンター系「C」の頻度は少なく全地方で最低でしたが、 人口比率が大きいため実人口数は他の地方より多いはずです。
中部は、広いため地方としてはまとめにくいのですが、特に大きく取り上げる特徴はないようですが、 あえて言えば「O3」の中で最も古い「O3」の頻度が高いことです。 「O3」の中でもオリジナルの黄河系が朝鮮半島に多く居住し「O3」の中でも新しい子亜型の進出で朝鮮半島を追い出されたようです。 ちなみに「O3」「O3a1」は黄河(華北)直系漢族、 「O3a2」は黄河系と長江(華南)系の交配系漢族です。
関西(近畿)は、まだ特殊なデータしかなくもっと調査が必要ですが、先に書いたように意外に縄文ハンター「C」が多い土地です。 しかし大阪、京都、奈良の大和朝廷族が攻略し占拠した 県のデータがそろうと圧倒的な「O3」地方になる可能性が極めて大です。また当サイトで以前卑弥呼は海洋性ハンターの呪術師で 支配者ではなかったかと書きましたが、 日本列島の全地方で最も「C」頻度の高い関西か四国が卑弥呼の居住地としては候補地でしょう。もしかすると入鹿、馬子、蝦夷などの 名前を持つ蘇我氏はこの「C」遺伝子集団だった 可能性は充分にあります。もしそうなら「C1a」だと思いますが...。
中国は、大国主神話もあり予想された葦原の国通り、弥生系が圧倒的に多い土地です。当然ながら「O3」武装侵攻軍の頭領神武や 物部のような別の「O3」侵攻軍も 「O2b」の抵抗にあい相当苦労して山陽道を進軍したはずです。そして「O2b」集団を征服するたびに「O3」支配者を常駐・在地化させ コントロールしてきたため、「O3」頻度も全地方最大なのでしょう。 その代わり縄文頻度は全地方最低です。最も日本らしくない地方です。遺伝子風土が異なるのです。
四国は、やや特異な遺伝子構成でY-DNA「D2」頻度は中国地方に次いで低いのですが「C1」はかなり高く海洋性ハンター漁民が 多く住みついた土地であることを示しています。 一方「O3」も多いと言うことは、大和朝廷族集団もしくは前後して日本列島にやってきた別の「O3」侵攻集団も瀬戸内海の南北を 要所として抑えながら東遷したことを伺わせます。 恐らく瀬戸内海側3県と太平洋側の高知県は、もっと詳細データが集まるとはっきりと区別できるのではないかと予想できます。 瀬戸内海側は「O3」頻度が高いはずです。
九州は、
縄文系Y-DNA「D2」「C1」は南から北上して青森までの行程の通過点だったことと、
同じ縄文系「C3a」はシベリア→朝鮮半島→九州とナウマン象など大型獣を追って南下してきた到達点、 (列島で最大の「C3a」頻度)です。
弥生系にとっては満州→朝鮮半島→九州と耕作適地を求めて南下してきた到達点だった がここから日本列島全土に拡大した。
プロト漢族系征服者「O3」にとっては、朝鮮半島を追い出された逃げ込み先で、旧唐書に言う「日本」 なる上陸拠点を確立した足がかりの土地のはずです。
沖縄は、
北琉球は国内データでは本土とほぼ同じ遺伝子構成であくまで日本列島の一部ですが、
南琉球はどうも違うようです。「O2b1」の土地のようですが詳細なGlobal論文を期待します。
琉球全土(特に北琉球)は南から北上してきた縄文系が住みついたためか、関東と並ぶ縄文頻度です。特にオーストラロイドと 判明した港川人は当然Y-DNA「C1a」だったはずです。 予想通り「C1a」頻度が高い土地です。南琉球はもし国内データ通りの「O2b1」地域なら「D2」との交配で「O2b」→「O2b1」に 分化した弥生系が九州から後代に南下し 「D2」を駆逐して住み着いたことになります。もし「O2b」なら朝鮮半島から一気に南下し定住したことになります。 いずれにせよ「D2」との交配より駆逐を選択したのかもしれません。
アイヌ民族は、当史観で繰り返し触れているように、縄文遺伝子Y-DNA「D2」が85%を占めるのですが、 本土が「O3」プロト漢族系に征服されたのに対し、アイヌは「C3c」古代ニヴフ族に征服され、 お互い被征服民族でしたが、本土では圧倒的な人口だったため先住民化せず、残念ながら北海道では支配者ともども 現代アイヌ民族として先住民化してしまいました。これが歴史のあやなのです。 同じY-DNA「D2」にも関わらず随分境遇が変わってしまいました。
以上
表紙に戻る
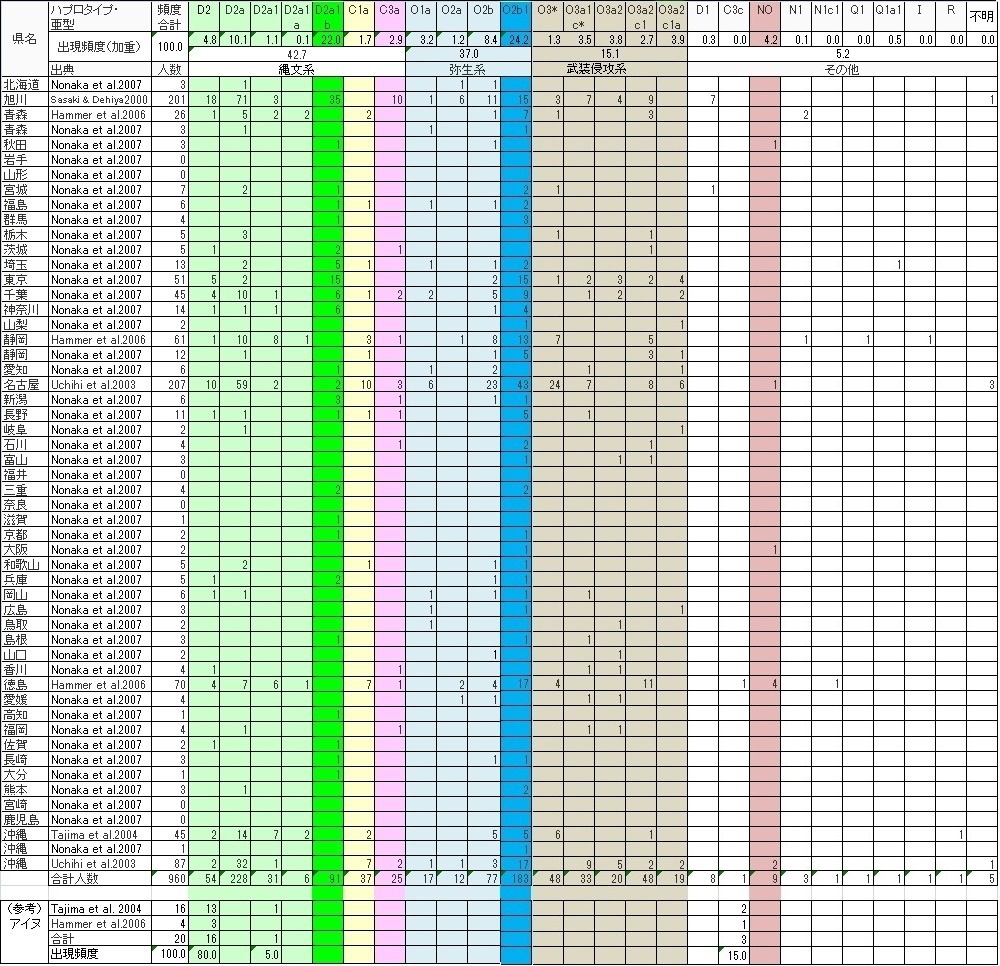
●県別データまとめ
当ガラパゴス史観はこれまで日本列島全土の遺伝子比率を使用し、琉球とアイヌのみを外出しにして論じてきました。 しかし、もし県別のデータがあれば違うアプローチの方法もあるのではないかと考え、これまで参考にしてきたGlobal論文を調べ、 その元になった県別のデータが一部入手できましたのでリストにして見ました。
先にご紹介したGlobal論文の中で県別の調査データが報告されていましたので、御紹介します。 このリストを見るとこの程度のサンプル数で民族論が論じられるのかかなり疑問になってきましたが、自分では調査できないので、 背伸びせずにガラパゴス的民族性を論じたいと思います。 とりあえず縄文系、弥生系、侵攻系と分けて頻度を出してみました。ご参考に!
最も利用されるNonaka et al.、Tajima et al.とHammer et al.の論文の他にNonakaが参考にしたUchihi et al.とSasaki & Dehiyaの 論文が県民と地方のY-DNAに触れていましたので、 それらの情報をまとめてリスト化しました。
日本人の合計データで見ると気がつかなかったのですが、県別(一部都市)で見ると1県あたりのサンプル数は極めて少ないのです。 これで何か物を言えるのだろうか?と極めて心配になります。
このリストで最も疑問なのは大阪で2サンプルしかないにもかかわらず、そのうちの1件をY-DNA「NO」が占めたため、 大阪は人口比率の大きいことから全体頻度にかなり影響をしています。 研究者は何故疑問をもたなかったのでしょうか?無作為抽出2人のうち1人が日本人の極少数派に当たるのは宝くじに当たるようなものです。 しかも関西圏は全てY-DNA「O3」が検出されていないのです。 征服者大和朝廷族のお膝元にもかかわらずです。
この論文の著者たちはこの宝くじ的な確立で出現したハプロタイプに何故再調査を行わなかったのでしょうか? 目的が日本人全体なので全国で見れば誤差範囲と考えたのでしょうか?
大阪は天皇陵古墳が存在する土地で大和朝廷族の本拠地でもあったにもかかわらず、です。 おまけに平城京の奈良県はサンプル数「0」です。著者はこの特異な場所であるはずの 関西圏で一体どんな人を対象にサンプルを抽出したのでしょうか?もし農村部や山間部を中心にサンプルを故意に抽出すれば 当然大和朝廷族のY-DNAは出てきません。大阪でうっかり抽出すると 近代の朝鮮半島系の出身者を調査する可能性は極めて大です。にもかかわずY-DNA「O3」の頻度が「0」という結果は わざとデータを操作した可能性が大きいのです。 関西は圧倒的な「O3」圏のはずだからです。
地域で最も平均的な頻度分布を持っているのは全国から人が集まっている東京と北海道だと思っていましたが、 東京はほぼ予想通りでした。しかし旭川は圧倒的な高縄文度の土地でした。 恐らく海外移民組も同じではないかと想像できますが、江戸時代まで抑圧されてきた貧民だった縄文の子孫が多く屯田兵や移民として 移住したからではないかと考えると納得できそうです。 海外の日系移民のY-DNA頻度を調査できると面白い結果が得られるかもしれませんね。
この結果は、県単位でサンプルを集めることの大変さと抽出のマジックを教えてくれます。目的が日本列島全体の遺伝子分布と 縄文-弥生の二重構造をあぶりだすことだったので、 限界があるのは仕方がありません。やはり研究者単位ではなく国が主導して調査をしないと高度な解釈は難しいようですね。
しかしそれでもこの県別調査で、大和朝廷族や武士団族に代表される征服者プロト漢族のY-DNA「O3」が日本列島にしっかりと 根付いていることと、 Y-DNA「C3c」に代表される東北アジア系騎馬民族、いわゆるツングース系騎馬民族王朝、は日本列島に全く乗り込んできていないことは 改めて明らかになりました。 恐らく朝鮮半島にいた中国王朝出先機関の漢族役人たちも朝鮮半島や周囲の先住民族たちと抗争するために騎馬民族的な風習を一 部取り入れていた可能性は否定できませんが、 あくまで漢族「O3」です。
縄文遺伝子Y-DNA「D2」と「C1」「C3」は日本列島で10000年もの長期間に渡り交配しお互いに子亜型に分化しています。 また4000年前頃から徐々にボートピープルとして 満州→朝鮮半島→日本列島に水田稲作の耕作適地を求めて南下してきた弥生遺伝子の呉系長江系Y-DNA「O2b」も縄文遺伝子「D2」との 数1000年の長期間にわたる交配で「D2」は更に新しい子亜型に分化し、 朝鮮半島と共通の「O2b」も日本独自の「O2b1」に分化しています。この結果は狭い日本列島で縄文-弥生集団が互いに 武装集団・戦闘集団ではないため、意外に敵対せず共存-交配をしてきたことを 如実に説明しています。
ところが、もともと侵攻者であった武装集団・戦闘集団のプロト漢族系Y-DNA「O3」は、日本列島上陸後1500年は 経っているにもかかわらず、日本列島独自の子亜型が全く存在せず、 中国大陸にあるそのままの子亜型のみしか検出されません。つまり支配階級はエリート層として縄文-弥生集団を差別し 交配を進めてこなかったことを如実に物語っています。 「O3」層が縄文弥生集団と交配をするようになったのは階級分化が崩れてきた戦後のことです。戦前はまだ身分の違いとかで めったに交配は起きてはいないのです。 今は世界の最先端を担う我が日本ですが、戦前までは「O3」の封建制がまだまだ根強く残っていたのです。
まだ良く理解できていないのはY-DNA「D1」です。集中的に見つかるのは旭川です。何故でしょうか? まさかチベットからの移民がいたとは考えられませんので、 「D2」が全く存在せず「D1」が数%存在する韓半島からの近代の移住者しか可能性はありません。 朝鮮半島の「D1」が或る時期に近代日本列島に移住し最終地に旭川を選んだのでしょうか。 もしこの論文の報告に間違いがなければ...ですが。2000年の論文なのでまだY-DNA同定技術が”稚拙”だった可能性は充分にありますが..。
残りのY-DNA遺伝子ハプロタイプは、朝鮮半島時代の中国出先機関(帯方郡、楽浪郡などで代表される)の「O3」支配下にあった 東北アジア系の民族でしょう。 特に「NO」「N」「Q」や「C3c」などは寒冷地適応し典型的なフラットフェースの一重瞼だったはずです。
以上
表紙に戻る