これでわかることは「C」系縄文人の近縁遺伝子が住むのは圧倒的にオセアニアが多いことがわかります。 「D」系はスンダランドから北上し東シナ海-黄海ランドからユーラシア大陸極東部に入りチベット族、羌族、Pumi族など チベット系諸部族となりましたが、 「C」系のかなりの部分がスンダランドをそのまま更に東進したようです。北上組みの中から更にベーリング海峡を渡り北米・中米まで 進出し分化したのが「C3b」、 シベリアに留まった集団の中から、その後寒冷地適応集団「C3c」以降が分化しました。
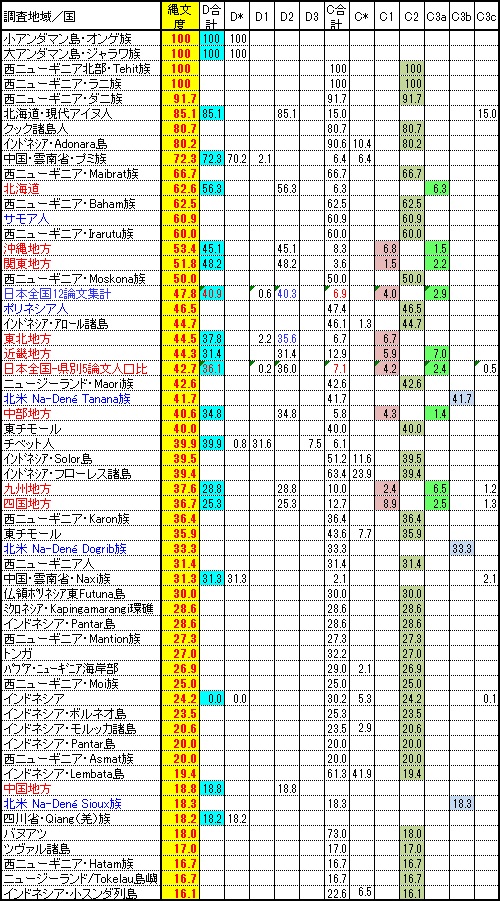
日本人でありながら、中国地方の縄文度の低さは目を見はるものがあります。 また長江文明の楚系と考えられるY-DNA「O1a」が突出して多いのも中国地方です、 大国主神話の特殊性や吉備の豪族一族などもしかすると楚系の文化・部族なのかもしれませんね。 ともかく中国地方は日本の中で特異な遺伝子地域であり、遺伝子的に異なる部族が住み独特の華南系文化を立ち上げていた 可能性が極めて高いのです。 大和朝廷族が朝鮮半島を追い出され流浪の武装集団として大和に向かった時に先住部族として立ちはだかった可能性は極めて高いと思います。 もう少し突っ込んで調べる価値はありそうです。
日本人のガラパゴス的民族性の核をなすのは、土着の縄文系ではなく武装侵攻系のY-DNA「O3」です。 楽浪郡、帯方郡など中華王朝の出先機関が牛耳っていた朝鮮半島での生存競争を勝ち抜くことができず、 流れ者武装集団として日本列島に逃げ込んできた戦闘集団の中で最後に勝ち残ったのが大和朝廷族となり、 敗れた集団は古事記などの言うその他天神集団となったようです。 最後に勝ち残った百済系大和朝廷族の親衛隊からのし上がった戦闘要員の中から平氏が生まれ、 高句麗から落ち伸びた戦闘集団の中から坂東武者が生まれ、新羅の落ち伸び花朗集団の中から源氏が生まれ、 このような朝鮮半島出自のY-DNA「O3」を中心とする武士団族が縄文-弥生交配集団が居住していた地方を制圧し 力を蓄えていったようです。
日本列島はこうして朝鮮半島発のY-DNA「O3」出自の大和朝廷族と武士団族という侵攻集団に征服されたのです。 そして漢語で読み書きをしていた征服者は先住の縄文-弥生集団をコントロールするために漢語を読み下しできるように 「かな・カナ」を開発し(ただし弥生系は長江文明の子孫なので発音は違っていても漢文・漢語は当然OKだったはずです)、 縄文系と弥生系の神話を自分たちの神話に付け加え、大和朝廷族が古代から日本列島を支配していたように記紀神話を 作り上げてきたのです。
しかし遺伝子調査が明確になるに従い部族・種族の移動が見えてきた現在、 記紀の神話の世界の中で、イザナミ・イザナギ神話は東ユーラシア大陸に最初に上陸した部族と考えられる羌族を核とする Y-DNA「D」族の伝承してきた 中国神話の伏羲・女?神話であることが見えてきました。
大国主神話は呉系か楚系の長江文明系神話で間違いないでしょう。過去の神話学は、遺伝子と部族・種族の動きが わからない時代の研究結果に基づきます。 新しい遺伝子拡散・移動情報を基に再構築する必要がありそうですね。
縄文人のコアを形成する子亜型Y-DNA「D2」の兄弟遺伝子ハプロタイプは、親亜型の「D*」と子亜型の「D1」と「D3」になります。 アンダマン諸島の「D*」のOnge族とJarawa族が縄文人祖形の直系子孫であることはいうまでもありません。
縄文文化で土器製作技術や狩猟などを担ってきたと考えられる技術戦闘集団は人口的にはマイノリティだった海のハンター Y-DNA「C1a」と陸のハンターY-DNA「C3a」です。 「D2」と行動を共にし南方から北上し琉球列島に港川人の遺跡を残し日本列島に上陸したプロトオーストラロイドと考えられる 「C1a」に最も近い弟遺伝子「C2」は、 スンダランドから北上せず東進しオーストラロイドとして現在ニューギニア各地域や他のオセアニア諸島に分布しています。 特にインドネシアに属する西ニューギニアのラニ族とTehit族は「C2」100%の生粋の縄文兄弟系部族です。
一方、陸組みの「C3a」は日本列島には上陸せず、一部の「D2」と共に韓半島からユーラシア大陸の内陸に入り大型動物を追って まだ温暖だったシベリヤに展開しましたが、 寒冷化に従い「D2」は当時陸橋だった間宮海峡を渡りサハリンに入り、やはり陸橋だった宗谷海峡から北海道に入り古代アイヌとなり、 一部は陸続きだった根室半島かカムチャッカ半島から陸続きだった千島に入り古代千島アイヌになったものと思われます。 一方シベリアまで「D2」と行動を共にしていた「C3a」は、「D2」と行動を共にせず寒冷化で南下したナウマン象などの大型獣を追って 南下し朝鮮半島から日本列島に入ったようです。 この時に南下せず新天地を求めてY-DNA「Q」や「R1」と共に当時陸続きだったベーリング海峡を渡り広大な大移動を決行したのが 「C3b」になります。 彼らは土器製作技術集団でもあったため北米南部から中米にかけて縄文土器似の土器を遺跡として残したようです。
一方、Y-DNA「C3a」のもう一つの弟遺伝子で、南下もせずアメリカ大陸にも渡らずシベリア大地に留まり寒冷地適応をしながら 東北アジアに定着したのが 後にツングース族やモンゴル族の先祖となったY-DNA「C3c」です。後に騎馬民族となった彼らは日本列島にも渡り騎馬民族制服王朝を 興したと言う学説が戦後大ブームになりました。 遺伝子解析の結果は明快で、日本列島本土に「C3c」の出現頻度は0に近く、騎馬民族制服王朝などはなかったことが明解に証明されました。
北海道を征服しオホーツク文化を立上げアイヌ人を熊祭りなど北方系文化に変えてしまったのは古代ニヴフ族になります。 残念ながら現代アイヌにどれくらい「D2」文化が残っているかは全く明らかにはなっていません、というより誰も研究していません。 熊祭りなどそれほど東北アジア文化に変えられてしまったのです。現代アイヌ自身も「D2」の風俗習慣はもはや全くわからないでしょう。 それでもY-DNAの約85%は今でもY-DNA「D2」であり縄文人「D2」と同根なのです。 遺伝子は風俗・文化・言語等環境で変わりやすい表面形質ではわからない不変の血統を明らかにするのです。
しかし本土の「D2」,「C1a」,「C3a」縄文系も、朝鮮半島での勢力争いで敗れ半島を追い出され武装勢力として日本列島に落ちのびてきた 武装侵攻集団Y-DNA「O3」の大和朝廷族や武士団族に征服され、結局北海道の「D2」と同じ”被征服者”の運命を歩むことになりましたが、 絶対人口が少なく古代ニヴフ文化に埋もれてしまった北海道「D2」文化に対し、本土では圧倒的多数派だった「D2」縄文集団は 現代まで基層の縄文文化を我々現代人が感じる縄文精神文化としていまだに維持できています。 3.11の時に世界から称賛された日本人の行動様式はこの縄文精神風土そのものなのです。土地に根付き勤勉で黙々と働き抜くのは 縄文気質なのです。
一方海洋性ハンターとして鯨を追ったりマグロを追いかけたり船を操り海の漁を展開する海洋性気質は同じ縄文人の 少数派Y-DNA「C1a」がオリジンです。 「C1a」縄文人は海に乗り出す海の民でもあるのです。
もう一方のY-DNA「C3a」は陸のハンターとして大陸を股に掛ける日本人の大陸的な気質のオリジンなのです。 「C3b」は北・中米大陸まで民族移動を行い、「C3c」ジンギスカンはユーラシア大陸を東西に横断しヨーロッパまで届くモンゴル帝国を築き ヨーロッパに低頻度ではありますがY-DNA「C3c」遺伝子を植え付けることに成功しました。
以上
表紙に戻る