2系統に分化したノルマン系Y-DNA「I1」とバルカン系Y-DNA「I2」ともども、ケルト系遺伝子「R1b」、スラブ・インドアーリアン系 遺伝子「R1a」との交配度も相当高く、またラテン系遺伝子「E1b1b」やセム系遺伝子「J」とも交配していて、ヨーロッパ人は遺伝子タイプが 複雑に交配しており、また言語も遺伝子頻度と必ずしも一致せずヨーロッパ人の過去の複雑な動きが見えてきます。 しかしさすがにスカンジナビアではラテン系遺伝子のY-DNA「E1b1b」の出現頻度は極めて低く交配は少ないようです、 アフリカに戻ったような遺伝子集団なのでスカンジナヴィアのような寒いところは苦手なのでしょう。
以上
表紙に戻る
Y-DNA「I」はヨーロッパの主要ハプロタイプで最も古いです、そして恐らくヨーロッパ内で小亜型に分化した唯一のハプロタイプ (非常に珍しい小亜型Y-DNA「C6」や他のハプロタイプの末端亜型等は別として)です。 それは、40000年前~30000年前にY-DNA「IJ」として中東からヨーロッパのどこかに到着したと思われ、 およそ25000年前にY-DNA「I」へ分化しました。言いかえれば、クロマニヨン人は最も恐らくはY-DNA「IJ」とY-DNA「I」に属しました。
Y-DNA「I1」小亜型は20000年前に分化したと推測され、旧石器時代後期と中石器時代のスカンジナビアで隔離されながら発展しました。 Y-DNA「I1」は少なくとも25のユニークな変異によって定義されます、それはこの血統が重大な人口減というボトルネック(絶滅危惧の)を 経験したことを示します。
このY-DNA「I1」に属する男性はすべて、10000年前~7000年前の間に生きていた一人の男性を先祖としています。
中石器時代の間、「pre-I1」および「I1」の人々は、連続するErtrbolle文化(紀元前5300年~3950年、下図のデンマークの辺り)、 Funnelbeaker文化(紀元前4000年~2700)およびPitted Ware(櫛目文)文化(紀元前3200年~2300年)の一部でした。
後の2つの文化は、農業の導入によって時々新石器時代の文化と見なされます。 しかしながら、ドイツの新石器時代の農民がスカンジナビアに少人数で遅く入り込みましたが、ライフスタイルは基本的に狩猟採集民の ままでした。このことが恐らく、スカンジナビアがヨーロッパで最も本質的な旧石器時代の祖先のうちの1人を保持できた理由です。
●Y-DNA「I1」はどのようにしてゲルマン系になったのでしょうか?
紀元前2800年から、東ヨーロッパからのインド・ヨーロッパ人の到着によって大規模な文化的・遺伝的な隆起がスカンジナヴィアを 襲いました。インド・ヨーロッパ人達は実際に新石器時代を経ずに一気に銅器時代および初期の青銅器時代をもたらしました。 スカンジナビアに到着した最初のインド・ヨーロッパ人は、現代のロシア、ベラルーシおよびポーランドの戦斧文化 (あるいは縄目文土器文化、または単葬墓文化)の人々でした。彼らは主にY-DNA「R1a」に属したと思われます。 これらの人々はスカンジナビアのY-DNA「I1」住民と同じ母方のmtDNA亜型mtDNA「U4」と「U5」を運びました。
スカンジナビアへの第2の主要なインド・ヨーロッパ人の移住は、Y-DNA「R1b」でした、このY-DNA亜型は中央ヨーロッパの Proto-ケルト-ゲルマン語の話し手の支流としてProto-ゲルマン語を導入したと思われます。Y-DNA「R1b」は、 恐らく後期Unetice(ウーニェチツェ)文化(紀元前2300年~1600年)の北方への拡大として現代のドイツからスカンジナビアに入りました。
最初にSigmund Feistによって1932年に提案されたゲルマン基板仮説によると、、proto-ゲルマン語はインド・ヨーロッパ語 (Y-DNA「R1b」と少し「R1a」)とpre-インド・ヨーロッパ語(在来の北欧Y-DNA「I1」)要素とのハイブリッド言語。この交雑は 青銅器時代の間に起こり、そして最初の本当のゲルマン文明である北欧-青銅器時代(紀元前1700年~500)の誕生をもたらしました。
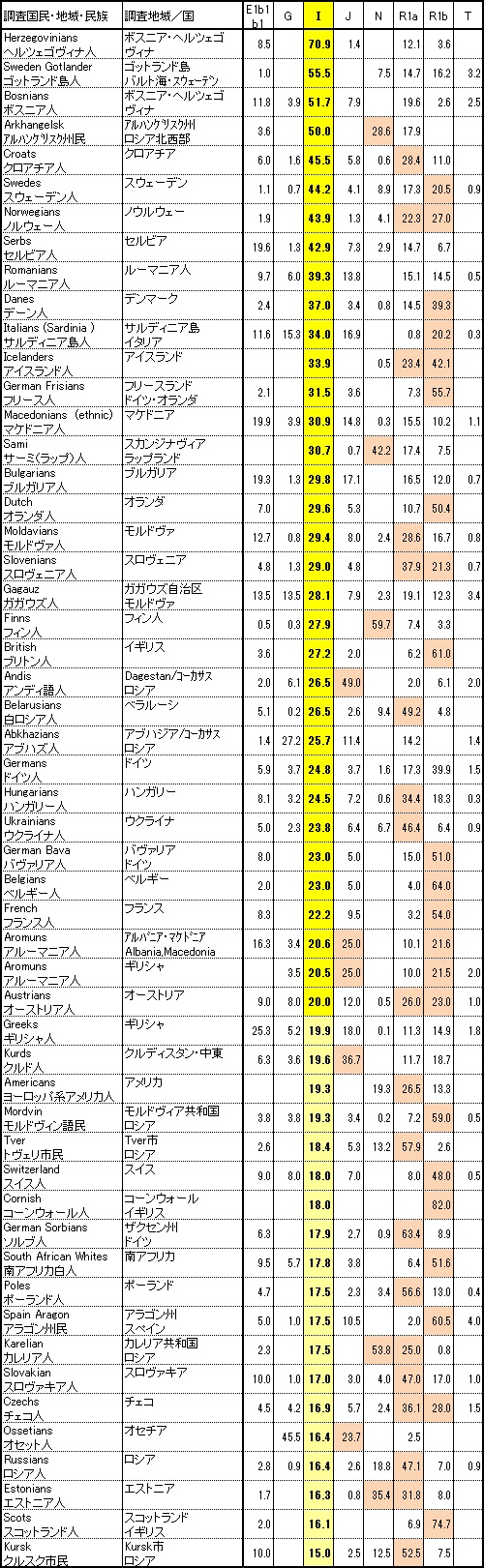
Y-DNA「I1」は北ヨーロッパで最も一般的なY-DNA「I」の子亜型です。それは、大部分スカンジナビアとフィンランドで見つけられ、 そこでは典型的で男性のY-DNAの35%%以上になります。ノルウェー人の植民に関係していて、Y-DNA「I1」は古代のゲルマン民族および ヴァイキングによって侵入されたすべての場所で見つかります。ゲルマン言語を話すヨーロッパの他の地域は、次の出現頻度を示しています。
つまり、ドイツ、オーストリア、低地地方、イングランドおよびスコットランドの低地地方はすべて、Y-DNA「I1」を10-20%を持っています。
以上
表紙に戻る
Y-DNA「l2」は、旧ユーゴスラビア、ルーマニア、ブルガリアおよびサルジニアで最も一般的なY-DNAの血統でありほとんどの スラブ諸国の主要Y-DNAです。Y-DNA「I2」の最大頻度は、ボスニア(55%)(ボスニアのクロアチア人の71%を含む)、サルジニア(39.5%)、 クロアチア(38%)、セルビア(33%)、モンテネグロ(31%)、ルーマニア(28%)、モルドバ(24%)、マケドニア(24%)、スロベニア(22%)、 ブルガリア(22%)、ベラルーシ(18.5%)、ハンガリー(18%)、スロバキア(17.5%)、ウクライナ(17.5%)およびアルバニア(13.5%)で観察されます。 ゲルマン民族諸国では5%から10%の頻度で見つかります。
Y-DNA「I2」(変異M438/P215/S31)は旧石器時代後期つまり最終氷河期最盛期の頃、約22000年前に起こったと思われます。 起源の地域は現在決定していません。それは、最終氷河期最盛期(LGM)で、局所的に種が生き残った場所/待避地の中の、 アナトリアかコーカサスの近くのどこかでしょう。
従って3つの仮説が可能です。
1.最初のシナリオは、Y-DNA「I2」がヨーロッパで始まったということです。
氷床が北へ後退し始めた20000年~12000年前頃、Y-DNA「I2」狩猟採集民はLMG退避
地から再度拡大し、西、中央、東ヨーロッパの広大な土地をを植民地化した。この
仮説では、Y-DNA「I2」はmtDNA「H1」,「H3」,「U5」および「V」に特に関係してい
るでしょう。
2.第2のシナリオでは、氷床が後退した時、Y-DNA「I2」は西アジアで始まり、ヨーロッパ
を植民地化しました。この仮説では、Y-DNA「I2」はほとんどmtDNA「J」および「T」
に関係しているでしょう。
3.3番目の最も有望でないシナリオでは、Y-DNA「l2」は西アジアで始まりましたが、新石
器時代までヨーロッパへ来ませんでした。
中東からヨーロッパへ新石器時代の農民および牛飼いの、Y-DNA「G2a」,「E1b1b」,「J」および「T」のような、 いくつかの独立した移住があったように思われます。それぞれのグループが1種あるいは恐らく2種のY-DNAハプロタイプだけをもたらしたのか、 あるいはほとんどの移住が既に多くのY-DNAハプロタイプが混じり合っていたのか、それは現在まだ明らかではありません。 この仮説では、Y-DNA「I2」はmtDNA「N1a」,「R」,「HV」,「H」(様々な子亜型),「J」,「T」,「K」および「X」に 関係しているかもしれません。
最初の2つのケースではY-DNA「I2」は、南東ヨーロッパ(M423)、中央ヨーロッパ(P214、L596)および地中海西部(M26)の新石器時代の 農民によって吸収されていたでしょう。Y-DNA「I2」の特定の子亜型の相対的な成功は農業の拡散にリンクされるように見えます。 南西のY-DNA「I2a1a」(変異M26)小亜型は、新石器時代のPrinted-Cardium陶磁器文化(紀元前5000年~1500年)の農民に吸収されました。 その子孫はほとんどが現代のサルデーニャ人およびバスク人で見つかります。
東のY-DNA「I2a1b」(変異M423)は、恐らくCucutani-Trypillian(ククテニ)文化(紀元前4800年~3000年)にリンクされます。 それは青銅器時代のインド・ヨーロッパ人の侵入の前にヨーロッパで最も高度な新石器時代の文化でした。
対照的に、中央のヨーロッパのY-DNA「I2」血統、北のY-DNA「I2」血統および西のY-DNA「I2」血統(変異L38、M223、L1286、 L1294およびL880のような)は、ほんの僅かの出現頻度で残存するのみです。
理由は、Y-DNA「I2」狩猟採集民が農業を採用したのが遅すぎたということであります、つまりインド・ヨーロッパ人が 中央、北・西のヨーロッパを占領したとき人口が少なかったからでしょう。
Y-DNA「I2a1」(変異P37.2)
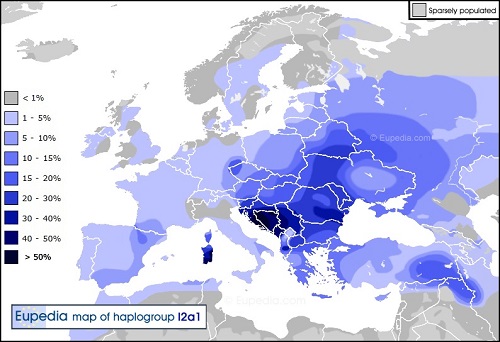
Y-DNA「l2a1」は、Y-DNA「I2」の断トツ最大の子亜型で、南東、南西と北西のヨーロッパで新石器時代の文化に 最も強くリンクされたものです。
Y-DNA「I2a1a」(変異M26)
Y-DNA「I2a1a」(変異M26、L158、L159.1/S169.1)は、2010年までI2a1と分類されていました。それはすべての西ヨーロッパに 分布しており、サルデーニャ人(37.5%)およびバスク人(5%)という2つの孤立した集団中で最大の頻度に達します。 Y-DNA「I2a1a」(変異M26)は、地理的に、英国の島、低地地方、フランス、ドイツ、スイス、サルディニア、シシリー、 イタリアの西海岸、イベリアおよびマグレブの地中海の西部海岸に制限されます。 Y-DNA「I2a1a」(変異M26)の子亜型のY-DNA「I2a1a1」(変異L160)はアイルランドにのみ制限されます。 Y-DNA「l2a1a」(変異M26)は、恐らく新石器時代・金石併用時代の期間に西ヨーロッパの巨石文化の主なY-DNA血統のうちの1つでした。
Y-DNA「I2a1」(変異P423)
Y-DNA「l2a1b」(変異M423、L1789)は2007年までI1b、および2008年から2010年までのI2a2として知られていました。 すべてのY-DNA「I2a1b」(変異M423)の主な小亜型の90%以上は、変異L621およびその小亜型の変異L147.2です。 他の変異は、変異L41.2(非常にまれ)および変異L161.1(ほとんどドイツおよび英国の島で見つかる)です。
Y-DNA「I2a1b3」(変異L621)
この小亜型は圧倒的にスラブの国々で見つかります。その最大の頻度は、ブルガリア人、ルーマニア、モルダビア、 西ウクライナおよびベラルーシと同様にバルカン系(ディナール系/Dinaric)スラブ人(スロベニア人、クロアチア人、 ボスニアのムスリム人、セルビア人、モンテネグロ人およびマケドニア人)でも観察されます。 さらに、それはアルバニア、ギリシャ、ハンガリー、スロバキア、ポーランドおよび南西のロシアでもより低頻度で共通に見られます。 Y-DNA「I2a1b3」1(変異L147.2)はバルカン半島のディナール(Dinaric)系に特徴的なため、Y-DNA「I2a-DIN」として知られています。
Y-DNA「I2a2」(変異P214)
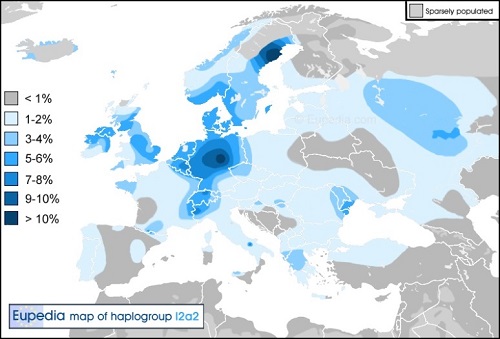
Y-DNA「I2a2」(変異S33/M436/P214、P216/S30、P217/S23、P218/S32、L35/S150、L37/S153、L181)は、2010年まで I2bとして知られていました。それは、巨石建築者(紀元前5000年~1200年)のような北西ヨーロッパのproto-ケルト-ゲルマン人に 関係しています。Y-DNA「I2a2」の持つ沢山の子亜型群は、「I2a2」が13000年の古さを持つことを示しています。
Y-DNA「l2a2」はすべての西ヨーロッパで見つけられますが、見たところでは、ドイツ北部でインド・ヨーロッパ人の侵入から うまく残存し、ラ・テーヌ(La Tene)文化のケルトの拡大(紀元前5世紀?1世紀)およびゲルマン民族の侵入(紀元3世紀?6世紀)の 両方によって再導入されました。 Y-DNA「I2a2」は現在、中央・北ドイツ(10-20%)をピークに、ベネルックス諸国(10-15%)や スウェーデン北部(10-15%)で見つかっています。またデンマーク、東イングランドおよびフランス北部でも住民の3?10%で見つかります。
ノルウェーにおいては、歴史上最もデ-ン人の影響が強かった南部を除いて、Y-DNA「I2a2」はまれです。
以上
表紙に戻る