Y-DNA「R1b」は、現在のいわゆるケルト系と呼ばれている民族に極めて頻度が高いためケルト系遺伝子と見なされていますが、 Y-DNA「I」に並ぶ現代欧州人の基層遺伝子の一つです。僻地のグレートブリテン島周辺に最高頻度が見事に残っています。 バスク人などイベリア半島にも同程度の頻度で残っています。ローマ時代は未開民族とみなされてきた辺境集団だったのです。
ノルマン民族のY-DNA「I」とやや異なる点は、同じように交配した遺伝子集団ですが、Y-DNA「R1b」は他の遺伝子の頻度が 低いことが解ります。特にウェールズ人やバスク人などは純粋な「R1b」に近いのです。 他の遺伝子集団との交配を嫌ってきた守旧的な集団なのでしょうが、独自の文化を守る意識が強かったのかもしれませんが、 純系を維持すると民族エネルギーは当然下がり、結局、辺境民族化してしまうのです。 人類は多様な遺伝子交配をすることでエネルギーを高めるように設計されているのです。 そして「R1b」もアフリカに進出していた一隊(Chadic語族)がいたことが確認できました。
ケルト系Y-DNA「R1b」が高頻度となる地域は西欧であることがよくわかります。 対して東欧はY-DNA「R1a」スラブ系が主役の地域なのです。
表紙に戻る
Eupediaに欧米人の「赤毛」はY-DNA「R1b」起源であることが書かれていましたので翻訳転載しますのでご参考に!。 赤毛のアンはケルト人の血統のようですね。Y-DNA「R1a」はブロンドの髪色が多かったようですが、「R1a」から分化した「R1b」は 赤毛の髪色が多いようなのです。つまりケルト系の多い地域は赤毛も多いのだそうです。
=========================
何が、赤毛を引き起こしますか?
赤毛は、16番染色体に位置する、メラノコルチン1レセプター(MC1R)の中の一連の突然変異によって引き起こされる劣勢遺伝の特徴形質です。 劣性形質として、髪が赤くなるには両親から遺伝子を継承しなければなりません。
従って、実際に赤毛を持っている人々より、はるかに多くの人々が赤毛のための変異を持っています。 スコットランドでは人口のおよそ13%は赤毛の人ですが、40%の住民が少なくとも1つの突然変異を持っています。
赤色でも、幾分明るい赤色、あるいはブロンドと混じりあった(「赤みがかったブロンド」)、幾分暗い赤色、赤褐色の髪 (それは赤みがかった色調を備えた茶髪)のように、 多くの種類の赤毛があります。
これは、一部の人々が1種類またはいくつかの考えられるMC1R突然変異の中の2~3種類しかもっていないからです。 髪の色の明るさは、皮膚と髪の両方の一般的な色素を形成する他の突然変異に結局依存します。
赤毛の事実
皮膚と髪の毛の色は、正常メラニン(eumelanin)とフェオメラニン(pheomelanin)の2つの異なる種類のメラニンによって引き起こされます。 最も一般的なものはeumelaninです。茶黒色のポリマーで、黒い色の髪と明るい色の皮膚の日焼けの原因になります。
Pheomelaninは、ピンク~赤の色調を持っており、唇、乳首および性器に存在しています。 MC1R遺伝子中の突然変異は、髪と皮膚に対してeumelaninより多くのpheomelanninを与えます、その結果、髪は赤くなり皮膚には そばかすを引き起こします。
赤毛の人は非常に色白な皮膚を持っており、大部分は非赤毛の人より常に色白い。 これは日光が希薄な北半球地方および非常に雨の多い国々で、より白い皮膚は日光の吸収を高めるので、身体によるビタミンDの生産には 大きな利点となります。 欠点は、赤毛は日焼けと皮膚がんの双方に対しより高いリスクを与えるということです。
研究は、赤毛の人々が熱の痛みにより敏感で、さらに他の髪の色を持つ人々より多量の麻酔薬を必要とすることを実証しました。 理由は、見たところでは、メラニン細胞刺激ホルモン(色素用の)とエンドルフィン(苦痛の取り除くホルモン)という少なくとも2つの 異なるホルモンに応答することができるホルモン受容体に、 赤毛の人が突然変異を持っているということです。
人々の知恵は、赤毛の人は、癇癪持ちで怒りっぽい(つまり短気)と、長いこと評してきました。
常染色体のDNA鑑定(例えば23andMeを備えた)を行うと、あなたがMC1R変異のうちのいくつかを持っているかどうかをチェック することができます。
赤毛はケルト-ゲルマン(Celto-German)の特徴なのか?
赤毛は長くケルトの人々に関係しています。 古代ギリシア人とローマ人の両者ともケルト人を赤毛の人と評しました。 ローマ人は、ゲルマン民族(少なくともローマ人がドイツ南部と西部で最も頻繁に遭遇した集団)にまで記述を広げました。 それは今日でもまだ該当します。
赤毛は北ヨーロッパと中央ヨーロッパのみのほとんど排他的な現象ですが、孤立したケースとして、さらに中東、中央アジア (タジク人の中に顕著に)で見つかります、 同様に(タリム・ミイラのうちの、中国北西部の新彊のタリム盆地で見つかるタリム・ミイラのいくつかに見つかりました。 ロシアのカザンとパーマの間のヴォルガ北部洗面器に住んでいるウラル語族のウドムルト人(高いY-DNA「R1b」頻度を持っている)は、 赤毛(10%以上)の高い発生率を持つ数少ない非西ヨーロッパ人です。 そうすると、これらの人々はみな何を共通に持ちますか? 確かに、ウドムルトとタジク人はケルト人でもゲルマン人でもありません。 そうです、これらの人々はみな単一のY-DNA「R1b」までさかのぼることができる共通の祖先を共有していることがわかりますね!。
赤毛はどの地域でより一般的ですか?
どこまで赤毛に含めるか、という定義の決め方に依存するため、赤毛を持つ人口の正確なパーセンテージを計算するのは難しいです。 例えば、髪の毛は赤くはないがあご髭が部分的に赤い男性を含めるべきか否か? 赤みがかったブロンドは赤毛もしくはブロンド?あるいは両方に数えられるべきですか?
定義に関係なく、赤毛の頻度はアイルランド(10~30%)とスコットランド(10~25%)で最も高いです。そして、 ウェールズ(10~15%)、コーンウォールと西イングランド、 (フランコ-ベルギー人が接している)ブルターニュ、それから西スイス、ユトランドと南西ノルウェーが続きます。 赤毛の出現率が1%未満になる地域との南部と東部の境界線は、スペイン北部、イタリア中部、オーストリア、ボヘミア西部、 ポーランド西部、バルト3国およびフィンランドです。
全体として、赤毛の配布は、古代のケルト人とゲルマン民族の世界に著しくよく一致します。 赤毛の最も高い発生頻度がケルト人のエリアで観察されることもまたは明白です。特に今日でもケルト語を話したり、あるいは 最近までケルト語を話してた地域で常に観察されます。 多くの人々の心に必然的に浮かぶ質問は次のとおりです: 赤毛の起源はケルト人もしくはゲルマン民族ですか。
ノルウェー南西部は多分赤毛の起源の手掛かりとなるでしょう。 ノルウェー南西部での黒い髪と赤い髪(ブロンドと対照的に)の両方の高い発生率は、Y-DNA「R1b1a2a1a2c」(変異L21)として 知られる父方の血統に高い頻度で同時に起ることが、 遺伝子系図のおかげで最近発見されました。 そしてそれはアイルランド北西部やスコットランド(Niall of the Nine Hostages:9人の 人質のニールと呼ばれる血統に)に典型な子亜型Y-DNA「R1b1a2a1a2c1a1a1」(変異M222)を含んでいるのです。 アイルランドとスコットランドのネイティヴ・ケルト人がViKingsによってノルウェー南西部へ(おそらく奴隷として)連れて行かれ、 そして、彼らがそこの赤毛の頻度を増やしたことは現在ほとんど間違いがないことです。
北緯45度線は赤毛になる突然変異が発生する自然的境界線か?
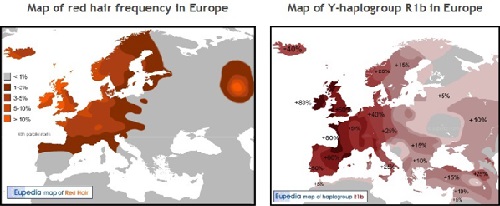
遺伝子系図学者にとって直ちに明白なことは、赤毛の地図がヨーロッパ北部と西部でY-DNA「R1b」の発生頻度と関連するということです。 明るいい色の皮膚と髪色の色素を含むいろいろな他の遺伝子を持っている人々の間で、赤毛はすぐ識別できるという単純な理由で、 それは南ヨーロッパではY-DNA「R1b」の パーセンテージとはまったく相関しません。 特に髪色が考慮される限り、地中海の人々はかなりより暗い色素(より高い正常メラニン)を持っており、赤毛対立遺伝子に赤毛を 発現させる機会をほとんど与えません。
赤みがかった色合いは、常に黒い髪によって隠されて、ダークブラウンの髪でもめったに見えません。 赤味(Rufosity)が劣性で、もし対立遺伝子が遺伝子プールでも分散し過ぎているなら、そして両親が対立遺伝子を持っている見込が 低すぎるなら、それは容易に隠れたままでいることができます。 さらに地中海域では、低緯度地域のより高い日光量とより強い紫外線が、より白い肌の赤毛の集団に対して潜在的に致命的な黒色腫を 引き起こしそうだったので、自然淘汰も地中海の住民から赤毛を次第に切り捨てました。
等しい緯度では、赤毛の頻度は、Y-DNA「R1b」血統の出現頻度と驚くほどよく相関します。 フランス中部 ・イタリア北部およびクロアチアを通る北緯45度の北は、赤毛頻度用の主な自然的境界であるように見えます。
北緯45度より低い緯度では、紫外線は非常に強くなるので、赤毛および非常に白い皮膚を持っていることはもはや利点ではありません。 北緯41度より低い緯度では、Y-DNA「R1b」の多い地域でさえも、赤毛の人は非常にまれになります。
北緯45度線はさらに、バターを基本とする料理の北欧文化とオリーブオイルを好む料理の南欧文化の間の従来の境界でもあります。 また、フランスでは、北緯45度線はフランス語の北のOil方言と南のOccitan方言の間の制限線です。
イタリア北部では北緯46度線が、イタリア語の話者からドイツ語の話者(南チロルの)を分けています。 北緯45度線が赤道と北極の正確に中間であるので、自然的境界は、恐らく一般に太陽および気候と非常に関係します。
はるか新石器時代にさかのぼっても、北緯45度線は、中央ヨーロッパのLinear Pottery(線形陶磁器)文化と地中海のCardium陶磁器文化を ザッと分けていました。 ヨーロッパの南北分割は、文化と農業のためだけではなく表現型と皮膚の色素沈着のためにも、それは充分可能であり得そうです。 新石器時代に戻ってみると、 ヨーロッパ南部の農業の拡大は主として中東からイベリア半島への地中海の海岸線に沿っった農民の移動に寄って実行されたのですが その時に、北のダニューブ川沿いの 農業の拡散は現地の中石器時代のヨーロッパ人自らによって成し遂げられました。彼らはテッサリアとアルバニア(Sesco文化)の農民と コンタクトすることで新石器時代の技術を 少ない範囲の交雑だけで手に入れました。
スラブ人、バルト3国人およびフィン人は、子亜型Y-DNA「R1a」、Y-DNA「N1c1」およびY-DNA「I1」の系統になります。 彼らの限られたY-DNA「R1b」系統は、MC1R突然変異がこれらの民族集団では非常に珍しいことを意味します。
そういうわけで、北西部のヨーロッパ人と同じ高緯度地域で生活し、明るい色の皮膚と髪の毛の色素にもかかわらず、部分的に ゲルマンの祖先をもつ数少ないポーランド人やチェコ人とは別に、 彼らのほとんど誰にも赤毛がありません。
どこで、赤毛は最初に起こりましたか?
特にネアンデルタール人が赤毛を持っていたことから、赤毛が旧石器時代のヨーロッパで発生したことが示唆されました。 ここまでテストされたただ一人のネアンデルタール人の標本(クロアチアから)は現代人(Arg307Glyとして知られていた問題の変異)の 赤毛の原因である同じMC1R変異を持っていませんでした。
しかし、ネアンデルタール人が600,000年の間ホモサピエンスと一緒に進化して、全ヨーロッパ、中東と中央アジアを横断し多数の 亜種を持った時から、ネアンデルタール人の1つの特定の亜種が MC1R突然変異をホモサピエンスに伝えた可能性を除外すれることはできません。
しかしながら、中石器時代と旧石器時代のヨーロッパ人に関連した唯2つの血統であるY-DNA「I」(例えばフィンランド、ボスニアや サルディニア))およびY-DNA「R1a」(東ヨーロッパ)の出現頻度が 最も高いヨーロッパの地域では、赤毛は存在しないかもしくは頻度が非常に低いため、これがヨーロッパで起こったとは思えません。 したがって、我々は驚きもしないが、どこか他で赤毛の発生地を探さなければなりません ― 青銅器時代の間にヨーロッパ中部と 西部に再植民したY-DNA「R1b」の人々に答があります。
Y-DNA[R1b]の起源は、複雑で、今日まで論争されています。 本Webの現在の著者は、Y-DNA「R1b」の、バルカン諸国、次に中央および西ヨーロッパへの青銅器時代の侵入の出発点として役立ち、 かつ北コーカサスおよびポントス大草原に移住が後続する、 中東の起源(反対する集団遺伝学者は極く少数ですが)に関する理論に賛成します。 この理論は、ウドムルト人、中央アジア人およびタリム・ミイラに存在する赤毛について説明する唯一の理論でもあります。
可能なネアンデルタール人とのリンク?
Y-DNA「R1b」は、恐らく上期旧石器時代のおよそ25,000年前にY-DNA「R1a」から分化したでしょう。 最も見込みのある場所は、現在のカスピ海の辺りで、最後の氷河時代が終わったあとロシア西部を覆っていた氷冠が融け、海になる だけだった中央アジアでした。 カスピ海が出来た後、これらの遊牧の狩猟採集民は、より緑でより豊かなカスピ海のコーカサス-アナトリアの側で、彼らは現地の 動物(例えば牛、ブタ、ヤギと羊)を飼育したかもしれません。
赤毛のための突然変異がネアンデルタール人から受け継がれたとするならば、おそらく現代のウズベキスタンまたはアナトリア西部/ メソポタミア地域に居住していた中央アジアの ネアンデルタール人からだったでしょう。 突然変異は、Y-DNA「R1b」部族によって受け継がれる前に、2000~3000年の間、若干の他(絶滅した?)の血統に、多分渡されたでしょう。 さもなければ、それは新石器時代(しかし、それより遅くない)と同じくらい後に、Y-DNA「R1b」集団の中で独立して発生することも できました。
赤毛とインド‐ヨーロッパ語族の移動
陶器類を自ら開発するかもしくはほとんど恐らく中東の隣人(特にY-DNA[G2a])から技術を得て、Y-DNA「R1b」部族 (そしてY-DNA[G2a3b1]の小さな少数派)の一部は、 彼らの家畜の群れのために草原の巨大な広がりを利用するために、コーカサスの向こうへ移住しました。 そこは、ユーラシアの大草原の在来のY-DNA「R1a」先住民に原インドヨーロッパ語族の文化が出現し広がって行ったであろう、 そしてY-DNA「R1b」集団が適度なレベルで 混じり合ったであろう場所です(今日どんな場所でも、Y-DNA「R1a」が優勢な東ヨーロッパからシベリアおよびインドまで、 Y-DNA「R1b」およびY-DNA「G2a」が少数派として 常に存在する理由なのです)。
起源前4000-3500年頃のヴォルガ-ウラル地域での馬の家畜化は、紀元前3300年前後の北コーカサスでの青銅器時代技術の出現と 結び付けられています。それはY-DNA「R1b」と Y-DNA「R1a」血統の素晴らしい拡大を導いたでしょう。そしてその冒険はこれらの原インド‐ヨーロッパ語族を、西はヨーロッパの 大西洋周辺へ、東はシベリアと北アメリカへ、 南はエジプトからインドまでの全ての道へと導きました。 紀元前3500年頃から、Y-DNA「R1b」の大部分は、金属資源に富んだバルカン地域へ、黒海沿岸に沿って西に移動しました。ここで それらは金石併用時代の「古いヨーロッパ」の先住民と混合しました。 シベリアと中央アジアに移動するY-DNA「R1a」血統に混じってついて行った少数のY-DNA「R1b」血統があったことが、Y-DNA「R1a」 血統が支配的な集団に赤毛がまれに出現する理由です。 (一部の部族が創始者効果のために彼らを失ったかもしれないが、通常まだ彼らの系統の中に少数派のY-DNA「R1b」を持っている)
考古学的な記録は、この継続したY-DNA「R1b」の一連の侵入がとても狂暴で、完全な破壊へと導いたものであることを、示します。 その当時までバルカン先住民とカルパチア先住民達の 繁栄する文明があったのです(彼らの子孫として、Y-DNA「I2a1b」とY-DNA「E1b1b1a」血統が生き残っています)。 Y-DNA「R1b」侵入者は妻や妾として先住民の女性を連れて行きました。そして新しい混合民族をつくりました。 言語は、先住民の古いヨーロッパの言語から借用語を採用し、結果として発展しました。 この新しい民族と言語の実体を原-イタリア-ケルト-ゲルマン語族と呼ぶことができるかもしれません。
ドナウ川の流域でのほぼ1千年後に(ババリアと同じくらいはるかに西へ)、彼ら(Y-DNA「R1b」)は紀元前2500年頃西ヨーロッパへと 西に向かう拡張を継続したでしょう。 実際、西方への拡大は、恐らくオーストリアとバイエルンの周辺のアルプス山脈の北に定住した、Y-DNA「R1b」の西端の一派によって 排他的に展開され、Unetice文化を発展させました。
多くのY-DNA「R1b」血統がバルカン地域に残りました。そこでは先住民とうまく混合し、そして次の1000年の間も後からこの地域に やって来た移民(とりわけY-DNA「J2」血統の農民、 ギリシャ、ローマ、ビザンティン帝国やオスマントルコ帝国の支配下での)ともうまく混合してゆきました。そして今日の地域における 非常に黒い髪の高い発生率のおかげで赤毛を失ってゆきました。 古代のギリシァの作家によると、赤毛はトラキア人の間で一般的でした。そして彼らは現在のブルガリア辺りに居住していました。 しかし今日では赤毛はほとんど完全に消えてしまいました。 しかしながら、赤毛対立遺伝子はローカルの遺伝子プール中で残存したかもしれませんが、明色の髪のための他の遺伝子の不足により 赤色を表現することができません。
赤い髪の原イタリア人、原ケルト人と原ゲルマン人は、中央ヨーロッパから連続した青銅時代のUnetice文化、Tumulus(塚)文化と Urnfield(骨壷葬地)文化が前進し拡大する間に、3つの語派に分裂しました。 原ゲルマン人は、小亜型Y-DNA「R1b1a2a1a1」(変異U106)を起源として始まり、現代のオーストリアからベネルクス一帯 (Low Countries)とドイツ北西部に移住したと考えられます。
彼らY-DNA「R1b」は、更にデンマーク、スウェーデン南部とノルウェー南部に(おそらく紀元前1200年頃から)拡大を続けます、 そこで、先住民のY-DNA「I1」やY-DNA「R1a」住民とうまく混じり合ったことで、古代ゲルマン民族文化は現れたのです。
現在のゲルマン民族の間の赤毛の頻度は、オランダ、ベルギー、ドイツ北西部およびユトランド半島(デンマ-ク)で最も高い、 つまりY-DNA「R1b」の出現頻度が最も高い地域です、 そして恐らく、鉄器時代の間にスカンディナヴィアから南方のドイツへと再拡大してきたY-DNA「R1b」および赤毛対立遺伝子の割合が 相当低いブロンド髪のY-DNA「R1a」および Y-DNA「I1」集団と交雑する前の、Y-DNA「R1b」によって最初に植民された地域でしょう。 赤毛はしたがって大陸の西ゲルマン語派と最も関係が深いのです。そして少なくともゴート人(Goths)とヴァンダル人のような スウェーデン起源のスカンディナヴィア人とゲルマン人とも関係があります。 これは、イングランド南部のアングロサクソンの定住が、なぜイングランド北東部のスカンジナビア人の定住より、高い赤毛の頻度を 持つかについても説明します。
イタリアの支系は、紀元前1300年前後頃にアルプス山脈を縦断し、イタリア半島の大部分、特にイタリア中部(ウンブリア人、ラテン人、 オスカン人)に植民しました。 彼らは恐らく子亜型Y-DNA「R1b1a2a1a2b」(変異U152)に主に属しました。
オリジナルのイタリック語派はケルト人やゲルマン人とちょうど同じくらいの赤毛を持っていただろうが、それらがエトルリア人の ように黒髪の隣人と交配したことで、赤毛を次第に失ったことはありそうです。 非インド‐ヨーロッパ語族(リグリア語、エトルリア人語、ラエティア語)が優位だった北イタリアへのガリアのケルト人の植民は、 したがって黒髪だったその地域に赤味(Rufosity)を増やしました。 最近では、イタリア中部で一般的な赤毛がイタリア北部でも一般的になっています。
ケルト人の分化は最大でかつ最も複雑です。
古代ギリシャ・ローマ時代でケルト話していた地域は、Y-DNA「R1b1a2a1a2」(変異S116/原-イタリア-ケルト語派のY-DNA)の いくつかの別個の子亜型に属する地方を包含しました。
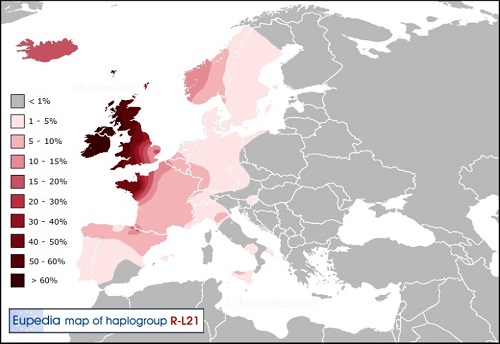
西ヨーロッパへのY-DNA「R1b」の最も初期の移住が、紀元前2100年前後の青銅器時代のフランス、ベルギー、イギリスおよび アイルランドへの拡散で起こったに違いありません。移住は、子亜型Y-DNA「R1b1a2a1a2c」(変異L21)と最も良く結びつきます。
2回目の移住は紀元前1800年前後にフランス南西部とイベリア半島へ起り、子亜型Y-DNA「R1b1a2a1a2a1」(変異Z196/バスク人に 特有]に結びつきます。 これらの2つの分化はその初期の分裂であったためケルト語と通常見なされますが、恐らく後の古代イタリア語と大陸ケルト語の 違いよりも明確ではなかったでしょう。
北西ケルト人の支系は、ゴイデル語(ゲール語)の祖先だったかもしれません、そして南西のものはイベリア半島の ケルティべリア人(Celtiberian)の祖先だったかもしれません。 両者はP-ケルト語派(ガリア語とブリトン諸語等が属する)に対立するものとしてのQ-ケルト語派に属し、 そしてそれはハルシュタット(Hallstatt)文化とラ・テーヌ(La Tene)文化およびY-DNA「R1b1a2a1a2b」(変異U152/古代イタリアの 支系と同じ子亜型)の拡張に関係しています。 今日では赤毛はケルト人の3つの支系のすべてで見つかり、それは子亜型Y-DNA「R1b1a2a1a2c」(変異L21)の中で最も一般的です。 理由は単純です、最北のケルト人支系(より高い地方でより有用な赤毛)であること、そしてイギリスとアイルランドのケルト人 の集団が最も純粋な原-ケルトの家系(非常に高頻度なY-DNA「R1b」)を保持してきたということです。
赤毛は、(中国のタクラマカン砂漠で発見された)約3000年前/紀元前1000年頃のタリム(盆地)・ミイラのうちの1体、 タータン着用の「Charchanマン」でも見つかりました(一緒に埋葬されていた3人の女性の内の一人はライトブラウンの髪でした)、 著者によると、この男性は中央ヨーロッパのケルト人の支流で現代のウイグル人の中のY-DNA「R1b」の存在に責任を負っています。 起源前2000年頃のタータンを着ていない初期のタリム・ミイラは、DNAテストの結果、Y-DNA「R1a」として識別され、 現代のY-DNA「R1a」が優勢な集団と同様に、赤毛を持っていませんでした。
以上
表紙に戻る
ヨーロッパの遺伝子の調査論文を調べてわかったことは、ゲルマン遺伝子のような単独遺伝子は存在しないと言うことです。 その代わり9種類程度の起源Y-DNA亜型が挙げられているようです。 Eupediaではこの中でウラル系Y-DNA「N1c1」と「N1c1a1」以外がまとめられていますので当ホームページで紹介しています。
何故ウラル系がまとめられていないのか?です。ロシアやバルト3国では必須のY-DNA「亜型なのですが?
Y-DNA「I1」 :ノルマン系
Y-DNA「I2」 :バルカン系
Y-DNA「R1a」 :スラブ系
Y-DNA「R1b」 :ケルト系
Y-DNA「E1b1b」 :ラテン系
Y-DNA「J2」 :メソポタミア農耕民系
Y-DNA「N1c1」 :ウラル系
Y-DNA「G2a」 :コーカサス系
Y-DNA「Q1a1a」 :フン系
どうやらゲルマン系と言うのは、Y-DNA「R1b」が核になったY-DNA「R1a」とY-DNA「I1」の交配集団のようです。 純粋に近いY-DNA「R1b」は残念ながらヨーロッパではケルト・バスクなど 辺境集団になってしまっています。
一方Y-DNA「I1」はノルマン系の単独遺伝子亜型であり、Y-DNA「R1a」もスラブ系の単独遺伝子亜型として現在に残っています。 ゲルマン系は3亜型が交配し形成された新しい民族集団であることが言えます。 つまり遺伝子が多様化するほど集団エネルギーは大きくなりますが、まさしくその通りの展開の好例がゲルマン系なのです。
そして同様にY-DNA「O3」を核に複数の遺伝子亜型が交配して構築された日本、中国、韓国がY-DNA「R1b」の覇権に 挑戦をしているのが現代の構図です。Y-DNA「R1b」もY-DNA「O3」も共に好戦的な遺伝子です。
和を持って尊しとする日本も、残念ながら大和朝廷族や武士族が持つこの好戦的なY-DNA「O3」がエグゼクティヴとして 支配階級を形成しているため日本人に非難される好戦的な1面を与えているのです。これが日本人の持つ2面性の要因なのです。
それにしてもヨーロッパでは子亜型、孫亜型までしっかりと分布が調査されていることがうらやましいですね。 日本だって縄文系Y-DNA「D2」の亜型群の地方分布をしっかりと調べれば、他の縄文系Y-DNA「C1a」と「C3a」との交雑、 弥生系Y-DNA「O2b」との交雑の歴史が少しは解明出来るのではないかと思います。
また関西圏のY-DNAを詳しく調査すると、大和朝廷族系Y-DNA「O3」、鯨漁等の近海漁労系と思われるY-DNA「C1a」と 修験道系と思われるY-DNA「C3a」の意外な分布がわかるのではないかと予想しています。
しかし武装侵攻系のY-DNA「O3」と本格交配が始まったのは明治維新後、それも多くは戦後なのでまだ「O3」との接触による 子亜型分化や孫亜型分化はまだ始まってはいないと思います。数百年もすれば新しい孫亜型群がかなり出現するようになるでしょう。
ではEupediaのY-DNA「R1b」の記事をご紹介します。訳が充分ではないので是非原文をお読みください。
====================================
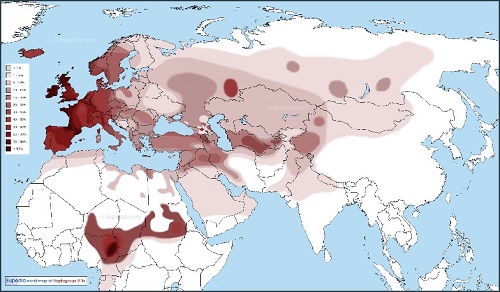
Y-DNA「R1b」は、アイルランド、スコットランドの高地、ウェールズ西部、フランスの大西洋の周辺、バスク地方および カタロニアで人口の80%以上に達する西ヨーロッパで最も一般的なY-DNAです。さらに、それはアナトリア、コーカサス周辺、 ロシア内の数ヵ所、および中央・南アジアでも一般的です。
ヨーロッパの大西洋および北海沿岸に加えて、イタリア北中部のポーの谷(70%以上)、アルメニア(35%)、ロシアのウラル山脈の バシキール人(50%)、トルクメニスタン(35%以上)、アフガニスタンのハザラ人(35%)、中国北西部のウイグル人(20%)および ネパールのネワール人(11%)等のホット・スポットを含んでいる。
Y-DNA「R1b1c」(変異V88;サハラ以南のアフリカに特有の子亜型)は、カメルーン北部で人口の60%~95%で見つかります。
旧世界のY-DNA「R1b」の分布図
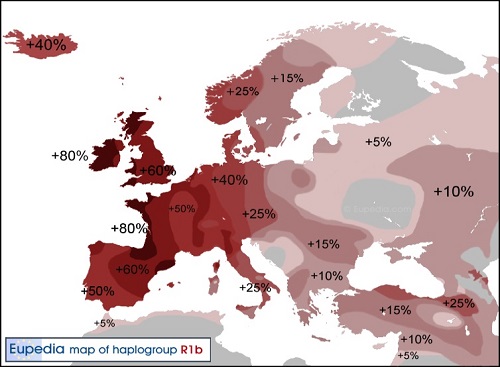
ヨーロッパのY-DNA「R1b」の分布図
Y-DNA「R1b」の主要な分化ツリー
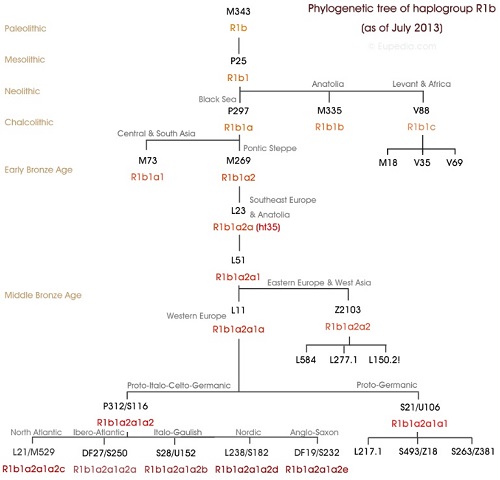
起源と歴史
●アナトリア起源かコーカサス起源か?
Y-DNA「R1b」の旧石器時代の起源は、今日まで完全に明らかではありません。
Y-DNA「R1b」の最も古い小亜型のうちのいくつかは、コーカサス周辺とイラン及び中央アジア南部で見つかります。 氷河時代に遊牧的なY-DNA「R1b」狩猟採集民が放浪することが出来た広大な地域です。
Y-DNA「R1*」および「R2*」は、中央アジア南部(カスピ海の低地とヒンドゥークシの間で)で始まったかもしれません。
Y-DNA「R1」の子亜型はそのときY-DNA「R1b」へ発展していたでしょう、そして最大氷河期最終期(およそ20000年前)頃の、 Y-DNA「R1a」が北のシベリアへ移動し空いた中東の北部あたりで、Y-DNA「R1b1」およびY-DNA「R1b1a」に分化したでしょう。
Y-DNA「R1b1a」は、新石器時代の間にアナトリア北部へ移動し、コーカサスを横切り、そこでY-DNA「R1b1a1」(変異M73)および Y-DNA「R1b1a2」(変異M269)へ分化したと推定されています。
現在、Y-DNA「R1b1c」(変異V88)へ発展した近東の痕跡は、レバノン人、イスラム教ドゥルーズ派およびユダヤ人の中に 低頻度で見つかります。
フェニキア人(今日のレバノンから来た)は、このY-DNA「R1b1c」をコロニー(特にサルジニア、マグレブ)へと広大しました。
Y-DNA「R1b1a2」(ヨーロッパで最も一般的な子亜型)およびY-DNA「R1b1a1」は、古代にインドヨーロッパ語が話されていた世界の すべての地域に存在することから証明されるように、インドヨーロッパ語の拡散に緊密に関係しています。大西洋岸からインド亜大陸まで、 ほとんど全てのヨーロッパ(フィンランドおよびボスニア・ヘルツェゴヴィナ以外の)、アナトリア、アルメニア、ロシアのヨーロッパ部分、 南シベリア、中央アジア周辺の多くの飛び地(ポケット)(特に顕著な新彊、トルクメニスタン、タジキスタン、アフガニスタン)および 忘れてはいけないのがイラン、パキスタン、インドおよびネパール、に存在しています。
Y-DNA「R1b」およびY-DNA「R1a」の歴史はお互い複雑に絡み合っています。
●北コーカサスおよびポントス-カスピ海の大草原 : インドヨーロッパ人のリンク
現代の言語学者は、原ヨーロッパ人の母国をポントス-カスピ海の大草原に置きました。明確な地理的・考古学的な地域は、 ドナウ川河口から東はウラル山脈へ、南は北コーカサスへ拡がった地域です。 ポントス-カスピ海の大草原の新石器時代、新石器時代と青銅器時代の間(Eneolithic)および初期の青銅器時代文化は、 塚(墓塚:クルガン)の下に死者を埋めるという長続きする習慣がその地域の文化として連続していているため、マリヤ・ギンブタス (Marija Gimbutas)によってクルガン文化(紀元前7000~2200)と呼ばれました。 墓塚(クルガン)式埋葬は起源前4000年紀から始まり、ほぼ間違いなく南コーカサスで起こったことが現在知られています。
馬は、起源前4000年頃のカスピ海の大草原の、恐らくドン川あるいはヴォルガの低地の周辺で、最初に家畜化され、 すぐに大草原文化を定義する要素になりました。しかしながら、Y-DNA「R1b」がその時の東部大草原の中に既にいたとは思えません。 したがって、馬の家畜化はY-DNA「R1a」に起因すると考えるべきです。
Y-DNA「R1b」がアナトリア東部から、いつポントス・カスピ海の大草原へ渡ったかは、まだ完全に明らかになってはいません。 それは新石器時代の間か新石器時代の直後に、もしくは両方で起きたかもしれませんでした。 Y-DNA「R1b」のコーカサス周辺での遺伝学的多様性は、大草原界に移動する前にY-DNA「R1b」がそこで既に発展していたたことを 否定するのは難しいです。
起源前5000年頃どころか起源前6000年頃のY-DNA「R1b」のアナトリアからの最初の移住が、大草原に羊を導入したということは 可能性があります。羊のウールは今日でもケルト人やゲルマン人(インド・ヨーロッパ語族のY-DNA「R1b」小亜型群)の衣類の 伝統に重要な役割を果たしています。
コーカサスを横切る別の移民が起源前3700年の直前に起こりました。つまりMaykop文化という世界初の青銅器時代の社会が、 北西コーカサスのどこからともなく現われました。 Maykopの起源はまだ不確かです、しかし考古学者は、アッシリアおよびイラン西部の同時代の金石併用時代の文化にリンクしました。
考古学は、さらにMaykop文化からポントスの大草原(Yamna文化がすぐ後の起源前3500年頃に発展)へのブロンズ製作と 墓塚タイプ埋葬の明確な拡散を示します。特に、クルガン(別名古墳)埋葬は古代のインド・ヨーロッパ語族の社会の支配的な 特徴になり、ケルト人、ローマ人、ゲルマン系部族、スキタイ人等によって広く使用されました。
Yamna時代(紀元前3500年~2500年)は、インド・ヨーロッパ語族の文化および社会の生成で最も重要なものです。
中東のY-DNA「R1b」の人々は、1000年以上恐らく2000年か3000年もの間、地元のY-DNA「R1a」の馬糧調達者や牛飼い達と うまく一緒に入り混じって住んでいました。
ポントス・カスピ海の大草原の至る所のY-DNA「R1a」とY-DNA「R1b」の人々の間の綿密な文化交流および相互作用は結果として、 言語学者が原インド・ヨーロッパ語(PIE)と呼んだ、 共通の自国語(新しい混成語)を生成しました。
原インド・ヨーロッパ語(PIE)の別の起源地域を探すことは無意味です。
言語の類似性は、一方では原インド・ヨーロッパ語族(PIE)と中東のコーカサス人とフルリ人の言語の間にも存在します。 そして一方ヴォルガ-ウラル地域のウラル語族の言語とも 類似性があります。それはポントスの大草原が完全な仲介の地域であることを意味します。
Yamna時代に、牛飼いと羊飼いはその食品とテントを輸送するためにワゴンを採用しました。それは結局インド・ヨーロッパ語族の 大移動に結びつく新しい移動の生活スタイルを生じさせ、 大草原へより深く移動することを可能にしました。
全部族がワゴンの助けを借りて移動したこの種の大量移動は、ジュリアス・シーザー時代のガリアや古代後期のゲルマン民族に おいてまだ一般的でした。
Yamna文化圏は一つの統一された文化ではありませんでした。
その南部は 、北西コーカサスまで黒海の北部沿岸に沿っった、東はカスピ海、シベリアおよびモンゴル(ユーラシアの大草原)まで 東方へと拡大する開いた大草原の地域でした。
その西部は、ドン川とドニエストル川(後のドナウ川)の間で、少数のY-DNA「R1a」集団(5-10%)を含む、Y-DNA「R1b」集団によって 最も密度高く植民されました。
その東部、ヴォルガ川流域からウラル山脈までは、Y-DNA「R1a」集団が多数でY-DNA「R1b」集団(バシキール人、トルクメン人、 ウイグル人およびハザラ人の中にその子孫は特に 見つけることができる)は基本的に少数派として居住していました。
Yamna文化圏の北部は、Y-DNA「R1a」集団によって占領された森林大草原でした。Y-DNA「R1b」(現代のロシア人およびベラルーシ人 の頻度から判断して、Y-DNA「R1b」の頻度は Y-DNA「R1a」の1/7~1/9程度の少数派に過ぎませんでした。
Yamna文化の西の支流はバルカン半島とギリシャへ、そして中央および西ヨーロッパに移動したでしょう、そして連続する波として 祖先の地アナトリアに戻ったでしょう (ヒッタイト人、フリュギア人、アルメニア人など)。
東の支流は、中央アジア、新彊、シベリアおよび南アジア(イラン、パキスタン、インド)へ移動したでしょう。
北の支流は、Corded Ware文化へ発展し、バルト海、ポーランド、ドイツおよびスカンジナビアに拡散したでしょう。
●Maykop文化、(Y-DNA「R1b」は大草原にリンクするのか?
北西コーカサスのMayKop文化(紀元前3700年~2500年)は、Yamna文化圏のある種の南へ文化の拡張です。
その地理学によりポントス・カスピ海の大草原文化の一部と一般には考えられませんでしたが、Mikhaylovka文化、Sredny Stoq文化 およびCkemi Oba文化等の同時の文化中の多数の セラミックス、金、銅、およびロンズ武器および宝石によって証明されるように、北コーカサスには大草原との密接なリンクがありました。
黒海北海岸と北コーカサスの間のリンクは、Maykop時代より古いです。
その先行文化者、Svobodnoe文化(紀元前4400年~3700年)にはSuvorovo-Novodanilovka文化および初期Sredny Stog文化へのリンクが 既にありました。 北コーカサス中のさらに古いNalchik植民(紀元前5000年~4500年)は、ヴォルガのKhvalynsk文化と似ていました。
これは、Y-DNA「R1b」が大草原のY-DNA「R1a」集団と相互に影響しあい混じり合い始めた時代かもしれません。
YamnaとMaykopの人々は両方とも、死者を膝を高くした仰臥位で置き、北東/南西軸の方向で墓塚埋葬しました。
墓には床に代赭石(red ochre)が敷かれ、人間と並んで家畜が埋められました。
さらに、2つの文化は共通の、乗馬、ワゴン、牛と羊に基づいた経済、銅/ブロンズ戦斧(ハンマー型斧と袖付斧の両方)および中子の ある短剣を使用しました。
実際、最も古いワゴンおよびブロンズ加工品が北コーカサスで見つかり、そこから大草原まで広がったように見えます。
MayKopは高度な青銅器時代文化でした、実際に非常に早く最初に金属製作を行いしたがって金属武器も開発しました。
世界で最も古い剣は後期MaykopのKlady墓塚31において見つかりました。
精巧な作りではありませんでしたが、その形はケルトの長い剣を思い出させます。
馬の骨と馬の描写が、初期のMaykopの墓には既に現れており、Maykop文化は大草原の集団、あるいは彼らと密接な関係にある 集団によって作られたかもしれないと示唆しています。
しかしながら、いくつかの遺跡の大草原文化と根本的に異なる文化的要素の存在は、Maykopが混成集団であることを意味する かもしれません。
DNA鑑定なしでは、混成集団の構成部族がアナトリアのY-DNA「R1b」グループおよびY-DNA「G2a」のコーカサス人グループの 2つの集団だったのか、Y-DNA「R1a」集団もそこに混じっていたのかは言うことができません。
これらの2つあるいは3つの民族は、異なる植民地で隣同士で住んでいたかもしれません。
インド・ヨーロッパ語族の移住のパターンに従う典型的なコーカサス人のY-DNA血統はY-DNA「G2a3b1」です。それはヨーロッパ、 中央アジアおよび南アジアの至る所で見つかります。
バルカン地域、ドナウ川流域および中央ヨーロッパでは、Y-DNA「G2a3b1」の頻度はY-DNA「R1b」の頻度に多少比例します。
Maykopの集団は、メソポタミアから大草原へ原始的な輪車両(ワゴン)の導入をしたと確信されている人々です。
それは、大草原の生活様式を革新し、後に起源前2000年ごろには戦争戦車の(馬に引かせた)開発に結びついたでしょう。
騎兵および戦車は、それらが速く移動し誰でも容易に打ち破ることを可能にしたことで、続いて起ったインド・ヨーロッパ語族の 移住の際に重大な役割をはたしました。
高度なブロンズ武器と海洋文化の結合された、黒海沿岸のインド・ヨーロッパ人の西の支流(Y-DNA「R1b」)は、謎の「海の民」の 重要な候補です。この「海の民」は起源前2000年頃に 地中海の東岸部を襲撃しました。
初期新石器時代のヨーロッパの文化分布
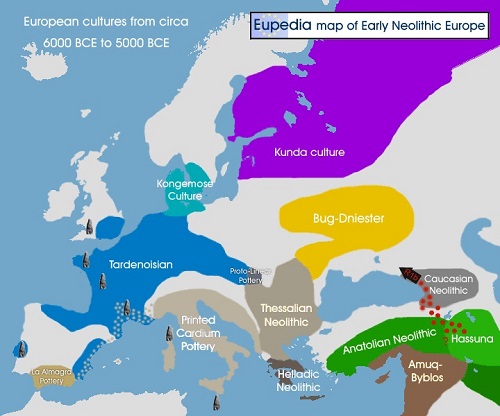
晩期新石器時代のヨーロッパの文化分布
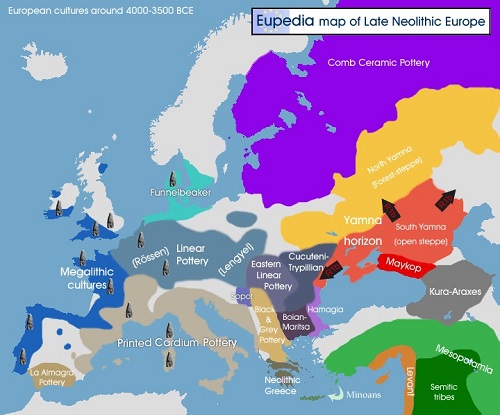
アナトリア中部のインド・ヨーロッパ語(IE)を話すヒッタイト人の興隆が、MaykopとYamnaの両文化の消滅の数世紀後に起こりました。
今日、Y-DNA「R1b」の最もインド・ヨーロッパ人らしい形質はアナトリアで見つかっています。それはY-DNA「R1b」(変異Z2103)の 子亜型Y-DNA「R1b1a2a2」に属します、そしてYamna/Maykopの集団が南東ヨーロッパに侵入した後にヒッタイト人がバルカン半島経由で アナトリアへ来たということにほとんど疑問を呈しません。
MaykopとYamnaの文化はSrubna文化(紀元前1600年~1200年)に受け継がれました。Y-DNA「R1b」民族が南東ヨーロッパとアナトリア に移動した後、空白地帯になった黒海沿岸へ Y-DNA「R1a1a」集団が大草原から前進してきたことが考えられます。
青銅器時代のヨーロッパの文化分布
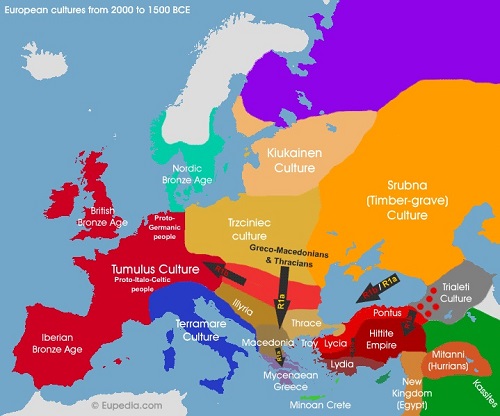
●Y-DNA「R1b」はどのようにして西ヨーロッパの先住Y-DNAハプロタイプに取って代わることができたのでしょうか?
最近までY-DNA「R1b」は、現在居住している地域でのその強い存在により、西ヨーロッパ起源と信じられていました。
その理論は、最後の氷河期のピーク時に旧石器時代のヨーロッパ人(クロマニヨン人)がフランスのカンタブリ地域に避難し、 氷床が後退した後にY-DNA「R1b」が中央および北ヨーロッパに再植民した、というものでした。
Y-DNA「R1b」の系統発生の調査は、より古いY-DNA「R1b」小亜型が中央アジアと中東で、また西ヨーロッパおよび北ヨーロッパに おいて最も若い小亜型が一貫して見つかったことで、このシナリオが可能ではないことを証明しました。
東から西へ、Y-DNA「R1b」集団の移住をトレースする明瞭な勾配がありました。
黒海沿岸から中央ヨーロッパへの主要な移住の時代は、さらにヨーロッパ(それは西ヨーロッパで青銅器時代文化の導入と一致する) のインド・ヨーロッパ語族の侵入およびItalo-セルティック語およびゲルマン語の増殖の時間枠と一致しました。
インド/ヨーロッパ語の移住が「大規模な侵入」だったのか、あるいは「言語の文化伝播」および「少人数の入来者」による技術の 広りだったのか、歴史家と考古学者は長い間議論をしました。
答えはおそらくどちらでもありません。
分散説の提案者は、Y-DNA「R1b」は西ヨーロッパに固有であり、Y-DNA「R1a」だけがインド・ヨーロッパ人を表わす、 と私たちに思わせるでしょう。問題は、Y-DNA「R」が中央アジアで発生したということです。そしてY-DNA「R2」は未だに中央アジアと 南アジアに限定され、Y-DNA「R1a」およびY-DNA「R1b」のより古い小亜型も中央アジアで見つかっている、と言うことなのです。
ヨーロッパのY-DNA「R1b」小亜型の分化の時代は青銅器時代と一致します。従って、Y-DNA「R1b」は青銅器時代以降のヨーロッパで、 ほとんど全ての先住・在来のY-DNAハプロタイプの血統と入れ替わったに違いありません。
しかしながら、大規模な移住および旧石器時代の集団のほとんど完全な絶滅は、ほとんど思い描かれた西欧人になりえない‥‥ すべてY-DNA「R1b」が支配的な地方であるにもかかわらず、アイルランド、オランダ、アキテーヌ(Aquitaine)あるいはポルトガルにおいて 全く異なっているように見える。常染色体のDNA研究は、西欧の住民が全く均質ではないことを確認しました。
多くの母の血統(mtDNA)が、さらに古代のDNA鑑定に基づいた旧石器時代の起源(例えばmtDNA「H1」、「H3」、「U5」あるいは 「V」)であるように見えます。
多くの人々が忘れるものは、patrilinealな血統がかなり速くとって代わられるために、大規模な脱出の必要もないということです。
理由は以下にあります。
1. 一夫多妻
女性と異なり、男性は作ることができる子どもの数に制限がありません。権力を持つ人
は典型的により多くの子どもがいます。これは、すべての未開社会(一夫多妻は多くの場
合、首領と王のための標準だった)で真実でした。
2. 地位と権力
青銅武器および馬を装備してインド・ヨーロッパ語族は、容易に新石器時代の農民を征
服し、ヨーロッパの最後の狩猟採集民はもっと容易に征服していったでしょう。彼らが
先住民の男を皆殺しにしないならば、新顔は平均より高い生殖の機会を持つため、地元
の王、族長と貴族の多数(青銅器時代のケルト人とゲルマン人は族長と小さな村落共同
体に住んでいました。そして、小さな種族の各々の一部が王によって率いられました。
)と共に新しい支配階級になったでしょう。
3. 男女比の不均衡
侵略軍には通常女性よりはるかに多くの男性がいます。したがって、男性は征服した住
民の中に女性を見つけなければなりません。戦争は男性によって行われます。また、敗
者はより多数の男性死傷者で苦しみ、勝者には利用できるより多くの女性が残されます
。
4.攻撃的な戦争
インド・ヨーロッパ語族は、勇気と武勇を強調する強い英雄的な規律を備えた好戦的な
集団でした。彼らの優れた技術(金属製武器、輪車両および軍馬)および生命に対する姿
勢は、彼らが金属製武器(つまり中東の文明以外の誰でも)を持った軍隊を組織していな
かったあらゆる部族・住民を虐殺することを可能にしていたでしょう。
5. 男子を妊娠する遺伝的な傾向
男性の体におけるY染色体の主な役割は精子を作ることです。亜型群(ハプログループ)
はY染色体を区別する変異に基づいて決定されます。それぞれの変異は精子生産および精
子運動性に影響を及ぼしやすいです。予備調査は、既に特定の亜型間のリンクと精子の
運動性が増加したか縮小したかを確立しました。精子の運動性がより高いほど、男の子
を妊娠するチャンスがより高くなります。Y-DNA「R1b」がより多くの男子の子孫を残
すことができたことは絶対にありえます。男子の出生のわずかに高い確率さえ、数千年
の間には蓄積効果で他のY-DNA血統に取って代わることにかなり寄与するでしょう。す
べてのY-DNA「R1b」小亜型にこの男子出生の偏りがあるとは限らないかもしれませ
ん。
同じ集団の中で見つかる他のハプログループ(亜型)に関して偏りが存在するだけです。 西ヨーロッパのかなり最近のY-DNA「R1b」 小亜型には、その地域の古いY-DNA亜型、特にY-DNA「I2」とY-DNA「E1b1b」(変異V13)と比較して有意な有利な条件があったことは、 非常にあり得ます。
父系(Y-DNA)の血統の置き換えは速く指数関数的な速度で進みます。
勝利者側のより高い地位は、彼らの子供たちのためにより大きな富、したがってより優れた栄養を保証します。 そして、成人期に達して、子供を作る可能性を増やします。負けた側の大部分の男性を殺して、彼らの女性(旧約聖書によって 証明されるように)を強姦するという青銅器時代的な古代的な行為・感性から見て、成人期男性比率の2:1は実は控え目な予想です。
そうなら、勝利を得た側のY-DNA血統はたった2、3世代程度で男性の大多数を占めるようになるでしょう。 たとえば、インド‐ヨーロッパ語族の最初の世代100人で各々息子2人が生き残るなら、先住民の生き残った男性200人は各々 息子1人のみで計300人なのに対して、インド‐ヨーロッパ語族の父の血統の数は第2世代では200人、第3世代では400人、第4世代では800人、 第5世代では1600人.....に増加してゆくでしょう。その間、先住民の男性血統は300人のまま停滞するでしょう。
そのようなシナリオに基づいて、Y-DNA「R1b」血統は急速に先住民の血統を圧倒していったでしょう。 たとえインド‐ヨーロッパ語族の征服者には先住民の男性よりわずかに多くの子供たちがいただけとしても、 Y-DNA「R1b」血統は2、3世紀以内に支配的になります。ケルトの文化は、首領と高貴の特権に終止符を打つローマ帝国の征服の前まで、 ヨーロッパ大陸で1000年以上の間続きました。これは、Y-DNA「R1b」血統が人口の50~80%に達するには十分な時間以上です。
現代のY-DNA「R1b」頻度は、ヨーロッパの大西洋周辺(最高頻度)から中央ヨーロッパと東ヨーロッパ(最低頻度)に向かって 頻度勾配があり、祖国のアナトリアで再び増大します。これは、粗野で好戦的なY-DNA「R1b」侵入者をヨーロッパ西部に進軍・移住 させておいて、農業が農業人口密度が高いヨーロッパ東部で、それから中央ヨーロッパへとしっかりと確立されたからにほぼ間違い ありません。
その上、他のCorded Ware文化(Y-DNA「R1a」)のインド・ヨーロッパ語族は、現代のロシアおよびウクライナの地域からドイツと スカンジナビアまで既に進んでいました。類似した技術と文化を共有した彼らの従兄弟遺伝子のY-DNA「R1a」集団と競争することは、 Y-DNA「R1b」集団にとっては難しいことだったでしょう。したがって、Pre-ケルト-ゲルマン語派のY-DNA「R1b」は更なる西に移住 しなければなりませんでした。最初にアルプス周辺に、そして当時人口のまばらな西ヨーロッパへとアルプスを越えました。
====================================
現在のY-DNA「R1b」の主要な子亜型群の分布図をご紹介します。
Y-DNA「R1b1a2a」(変異L23):バルカン、ギリシャ、アナトリアとアッシリア系子亜型
現ヨーロッパY-DNA{R1b1a2a****」の親子亜型、アナトリア、メソポタミアを中心にほぼヨーロッパ全土に拡がっている。
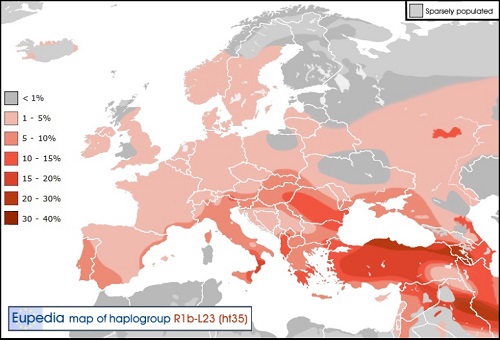
Y-DNA「R1a1a2a1a1」(変異S21):ゲルマン系子亜型
ドイツ、オランダ、ベルギー、オーストリー、イングランド、デンマーク、スウェーデン南部、ノルウェー南部等いわゆるゲルマン 民族圏。イギリスでは先住民ケルトの土地にゲルマンの一派アングロ・サクソンが侵入し、ケルトをスコットランド、ウェールズと アイルランドに追いやった歴史が見事に理解できる。
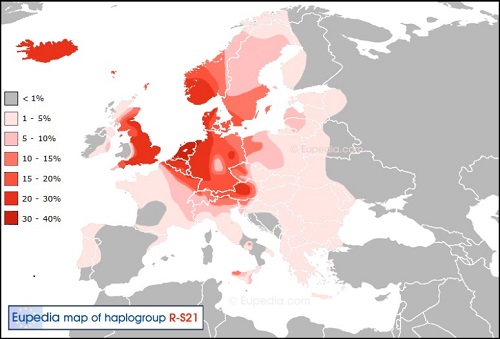
Y-DNA「R1a1a2a1a2a」(変異DF27):ガスコーニュ、イベリア半島系子亜型
ガスコーニュ語、アラン語を話す集団が居住する地域で、カタロニア州やバスク地方がスペインから分離独立を叫ぶ理由はこの 遺伝子に基づく文化・歴史の違いが大きな要因である。 ではフランスのガスコーニュ地方はどうするのだろうか?フランスの一部になりきっているようで特に分離独立の動きはなさそうです。
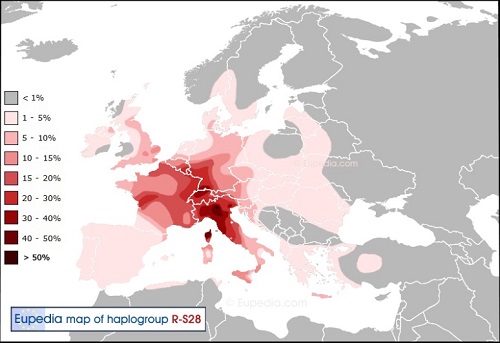
Y-DNA「R1a1a2a1a2b」(変異S28):イタリア-ケルト系子亜型
Y-DNA「R1b」の子亜型群の中でアフロ-アジア言語の文法が部分的に採用されている子亜型、近東の農民だったY-DNA「E1b1b」、 「G2a」、「J2」と「T」との混合が強かったためと考えられている。イタリアのトスカーナ地方が中心のようだ。
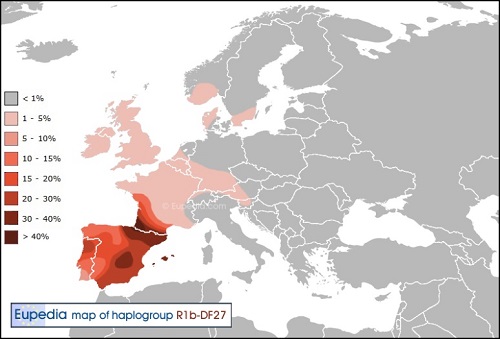
Y-DNA「R1a1a2a1a2c」(変異S28):大西洋-ケルト系子亜型
日本人がケルトと思っているのは、この遺伝子が核の文化になります。スコットランド、ウェールズ、アイルランド、ブルターニュが 居住地の、典型的な辺境Y-DNA「R1b」です。 頻度が80%を越える地域もあり、ガスコーニュ、バスクと並び純粋なケルト系Y-DNA「R1b」が残っている地域になります。
本来のケルト系はスラブ系と共に音楽や芸術等文化的な優れた感性を備えた優しい気質を持っています。
ところがゲルマン系は同じY-DNA「R1b」を核にしながらも、先住のY-DNA「I」や従兄弟遺伝子のY-DNA「R1a」と交雑し 更にY-DNA「G2a」、「E1b1b」や「T」とも交雑することで集団エネルギーを圧倒的に高め、恐ろしいほど好戦的に変貌したため ヨーロッパを制圧できたのです。
極めて明らかなことは、国・国民が先進的になるには純系民族では無理なのです。辺境民化してしまいエネルギーが低すぎるのですが、 競う共存遺伝子の種類が多ければ多いほど集団エネルギーが高くなり、国の活性度が上がり、覇権に向かうのです。
アジアの中で唯一近代化に成功した日本は縄文系-弥生系(長江文明系)-武装侵攻系(黄河文明系)が交雑し、 武装侵攻系が核になり集団エネルギーを一気に高め、一時はジャパンアズNo1と覇権を握るかもしれないほどの勢いを手に入れました。
しかし日本が高止まりしてしまった間に中国が、日本以上の複雑な遺伝子ミックスにより集団エネルギーを高め、近代化らしきものに成功し 対外的には日本に取って代わりアジアの覇権を握ったように見えるレベルに達しました。しかし国民一人当たりの生産性があまりにも低く、 日本の1/10程度しかなく真の覇者には恐らく永久になれないでしょう。
その中国も恐らく近いうちに日本同様高止まりするでしょうが、東アジアには日本、中国に代わる国はもはや存在しません。
南アジアのインドはロシアのスラブ系と同じインド・ヨーロッパ系遺伝子民族ですが、中国同様あまりに国民一人あたりの生産性が低すぎ 覇権を握るのはまず無理でしょう。
当分の間はY-DNA「R1b」のアメリカとY-DNA「O2」(旧「O3」)の中国が覇権争いを続けるでしょうが、中国も日本もアメリカに 対する輸出で生産性を上げてきたので、アメリカにとって代わることは逆に自滅に向かうためまず不可能でしょう。
非常に残念ですが、現代世界の構図は、世界の警察官であり輸入超大国のアメリカが太陽として中心に存在し、その周りに各国が 衛星のように回っているだけなのです。水星、金星、地球、火星ぐらいまではEU諸国、木星が日本、土星が中国という感じでしょう。
以上
表紙に戻る