ヨーロッパの3大遺伝子タイプに続くのはY-DNA「N」です。しかしXX民族遺伝子と呼ばれる総称はありません。 この遺伝子を持っている民族は全てシベリヤ大地に居住する民族とそのまま西に平行移動した結果ヨーロッパ北部の似た気候の土地に 住み着いた集団です。 したがってあえて言えばシベリア系民族です。寒冷地適応を成し遂げた民族なのです。極東シベリヤほど高頻度なY-DNA「N」が残るため この辺り、 特にヤクート族の居住するヤクーティアが発祥の地のようです。西に進むに従い他の遺伝子との交配が進んでいます。 言語的にはテュルク語系、ウラル・フィン語系とバルチック系のほぼ3言語に集約されます。前回のスラブ度チェックにバルト3国が全て リストアップされていましたが、 シベリア度チェックでも3国全てリストに見える見事な交配度なのです。そしてスラブ度では出てこなかったフィン語系の民族が顔を出します。 ヨーロッパ3大遺伝子と異なるのはY-DNA「P」、「Q」や「D」との交配がある民族もでてくることです。
ユピク人はエスキモー系です。シベリア系の多くは西に進みましたが、東のアラスカまで進んだ少ない例がユピク人です。移動時期は アリューシャン列島が陸橋で繋がっていた頃でしょう。
以上
表紙に戻る
Eupediaには今までY-DNA「N」の詳細が無かったのですが、やっと詳細説明が記載されたので翻訳転載します。
Y-DNA「N」はユーラシア大陸を10000〜12000年前頃に東から西へ横断大移動を行った遺伝子のようですが、有史前の出来ごとになると 該当する民族移動を探すのは難しいようですね。
欧米の王朝先祖調査FTDNAプロジェクトによると、ロシアのリューリク王朝のウラジミールⅡ世(1053-1125)はY-DNA「N1c1」だった らしいく、従兄弟のOlegⅠ世はY-DNA「R1a1a」だったらしい。 しかもこのリューリックのY-DNA「N1c1」(変異:DYS390=23)子亜型は現在ロシアのウラル語族内ではなくスカンジナヴィアで見つかるそうだ。 歴史はなかなか複雑でまさしく小説より奇なりのようだ。
以下、Eupediaの記事の翻訳です。
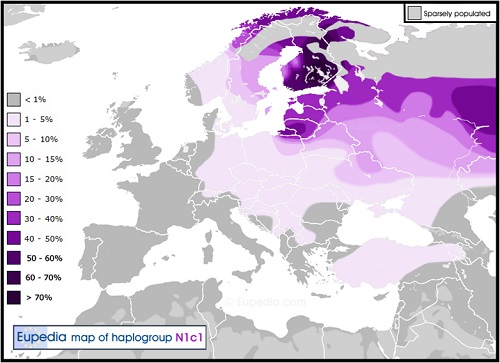
Y-DNA「N1c」は、主として東ヨーロッパの北部、特にフィンランド(61%)、ラップランド(53%)、エストニア(34%)、ラトビア(38%)、 リトアニア(42%)および北ロシア(30%)で高頻度に分布し、 さらにロシア中央部(15%)、ベラルーシ(10%)、東ウクライナ(9%)、スウェーデン(7%)、ポーランド(4%)およびトルコ(4%)でも低い頻度で 見つかります。
Y-DNA「N1c」はヴォルガ-ウラル地域のウドムルト人(67%)、Komi人(51%)、マリ人(50%)およびモルドヴァ人(20%)を含むウラル語族の 集団で顕著にみられるが、隣人のテュルク語族のチュバシ人(28%)、 ヴォルガ・タタール人(21%)およびバシキール人(17%)でも顕著で、同様に南ロシアのノガイ人(9%)にも見られる。
ヨーロッパのY-DNA「N1c」は、Y-DNA「N」のユーラシア西端の到達範囲を表わしていますが、全極東(中国、朝鮮、日本)、モンゴル およびシベリア(特にウラル語を話す北シベリア住民の間で)の至る所で見つかります。
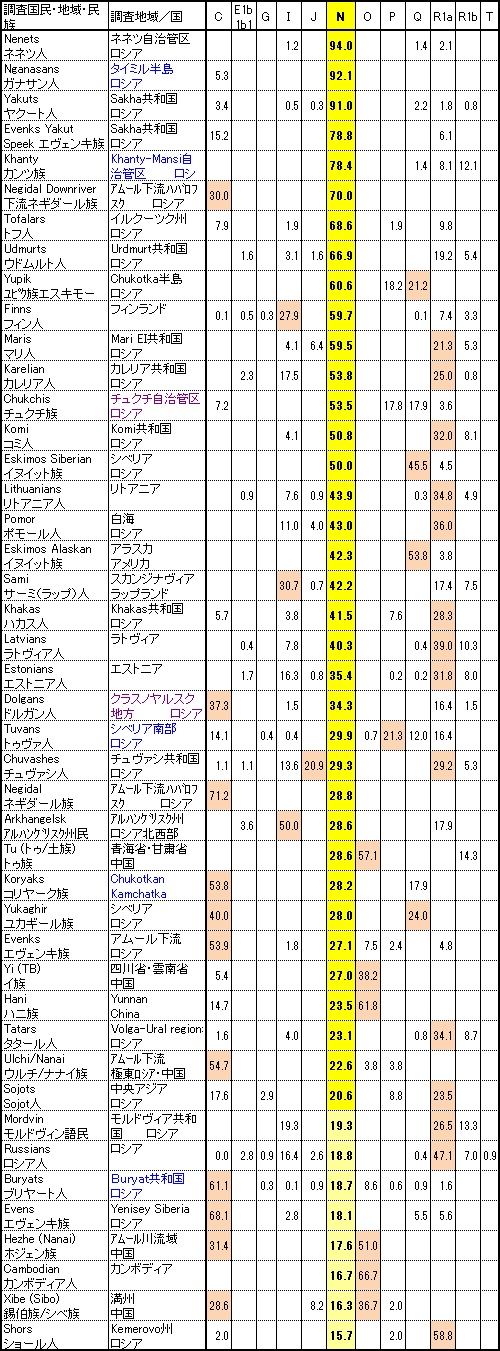
Y-DNA「N1c」は、中央シベリア北部の2つのウラル語族、ネネツ(Nenet)族とガナサン(Nganasan)族で最大頻度の95%近くに達します。 また中央シベリア東部のサハ(Sakha)共和国(Yakutia)に主に居住するテュルク語族のヤクート族では90%になります。
ヨーロッパのY-DNA「N1c」の分布
Y-DNA「N」は東アジアのY-DNA「NO」の子孫です。
それは、およそ15,000-20,000年前にインドシナか中国南部で始まったと考えられます。
Y-DNA「N1*」およびY-DNA「N1c」は両者とも、中国北東部の西遼河の渓谷の新石器時代〜青銅器時代(4500-700 BCE)の遺跡から 37%(70サンプル中26サンプル)の高頻度で見つかります(Yinqiu et al. 2013)。
新石器時代のサンプルの中でも、Y-DNA「N1」が紅山(Hongshan)文化(4700-2900 BCE)と小河沿(Xiaoheyan)文化(3000-2200 BCE)から 得られたサンプルの3分の2の頻度を占めると言うことは、Y-DNA「N1」集団は中国北東部および恐らくモンゴルとシベリアでも新石器時代の ライフスタイルの拡散に主な役割を果たしてきたと推測できます。
======================================
?ガラパゴス史観註
最近の古代中国史では中国4大古代文明は、長江文明、黄河文明、遼河文明および四川文明ですが、それぞれ主役のY-DNA亜型は異なり、
・長江文明はY-DNA「O1」と「O2」が主役、
・黄河文明はY-DNA「O3」、
・四川文明はY-DNA「D1」と考えられる、縄文人の中心の「D2」の兄弟亜型、
・遼河文明はY-DNA「N1」が主役と考えられます。
Y-DNA「N」とY-DNA「O」はY-DNA「NO」から分化し、「O」は長江と黄河流域で発展したのに対し「N」は遼河流域で発展し結果的 でしょうが棲み分けしていたようです。「D」は中国大陸/中原の 最初の民族のはずでしたが「O」、特に「O3」に追われ「O3」が好まない標高の高い高原部に逃げ込み後の羌族やチベット族を形成 したようです。
ところが一大古代文明を築いたはずのY-DNA「N1」も結局、恐らく「O3」によって遼河からシベリアに追われ古住シベリア民として ひっそりと生きることになったようです。
残った「O1」,「O2」も大部分は「O3」集団に取り込まれ長江文明系黄河系遺伝子集団に変質し漢民族化したのですが、同化を 拒んだ集団のうち、「O1」(恐らく楚民)は海南島や台湾経由で東シナ海や 南シナ海に逃げ諸島の先住民化し今に至っています。
「O2」の中で「O2a」(恐らく越民)は南に逃げ今日のヴェトナムや東南アジア民となり、更に西に進みインド亜大陸でモンゴル系 先住民となり更に南下してドラヴィダ民族の中で 水田稲作農耕民として生き残り、特にタミール人に水田稲作農耕文化の語彙を多く残し、大野教授が日本語のタミール語起源説を 発表したのは有名である。
一方北に向かった「O2b」(恐らく呉民)は満州辺りで「O2a」から「O2b」に分化し、水田稲作の適地を求めて南下し韓半島南部で 稲作を発展させ現在でも韓国民の30%以上のY-DNA頻度を もたらすほど定着している(これが韓国南部が日本と極めて近い理由の大きな部分)。
ところが「O2b」は更に水田稲作適地を求めて対馬海峡を南下し恐らくボートピープルのように小集団で繰り返し日本列島に移住した ものと考えられ、先住民のY-DNA「D2」縄文人に恐らく 敵対せず入り込み交配を重ね「O2b1」に分化し、水田稲作農耕文化を日本列島に定着させたものと考えられます。「D2」系縄文人の 強い敵対がなかったため青森まで稲作が早いスピードで 北上できたものと考えられます。
「O2b」が長江文明系の呉民出自であったため後の漢字の導入に際し受け入れに抵抗がなく、導入時に「O3」の大和朝廷族が採用した 唐音に加え「O2b」の持っていた呉音が日本語に残る結果に なったものと思われます。
日本列島へのY-DNA「N1c」や「NO」、「Q1」などの移動は恐らく、朝鮮半島の中国王朝の出先機関で生き残り抗争に負けて 追い出された武装侵攻「O3」集団の大和朝廷族や物部などの他の 天孫族や武士団族の配下や郎党として渡ってきたのでしょう。ただし縄文晩期に見つかる陸稲農耕は漢民族が形成される前の古い「O3」が 持ち込んだと考えるのが合理的ですが(「O3」で最も古い 「O3*」が日本人に見つかる)、その際に「NO」,「N1」や「Q1」が渡来した可能性もあります。
======================================
ヨーロッパで見つけられるY-DNA「N1c1」子亜型は、12,000年前に南シベリアを横断し、10,000年前にヨーロッパ北東部に拡散したと 考えられます。
それはクンダ(Kunda)文化(8000-5000 BCE)および後の櫛目文土器(Comb Ceramic)文化(4200-2000 BCE)に関係し、 フィン人やプレ-バルト人へ発展しました。
インドヨーロッパ語族の縄目文土器(Corded Ware)文化(3200-1800 BCE)は、紀元前2,500年前頃からバルト海沿岸地域および フィンランド南部に進出してきました。
2つの文化の混合はハイブリッドKiukainen文化(2300-1500 BCE)を生じさせました。
現代のバルト人は、ウラル語族とスラブ語族の文化の合併に起因する、Y-DNA「N1c1」とY-DNA「R1a」のほぼ等しい割合を 持っています。
低頻度のY-DNA「N1c1」は、すべてのスラブ語族やスカンジナビアの集団に見つかり、同様に北西部を除いたドイツの大部分 でも見つかります。
その起源は現時点ではまだ不確かですが、最も恐らくはロシア、ベラルーシおよびウクライナからドイツ東部に向かった鉄器時代及び 中世早期のプロト-スラヴ種族の拡散によるものでしょう。
スカンジナビア人のY-DNA「N1c1」は次の3つのオリジンの可能性があります。:
1. 鉄器時代以来のスカンジナビア人/ドイツ人の集団による北部サーミ(Sami)人集団の進歩的な同化による。
2. 最近の2000年の間に起ったドイツおよびポーランドからの移住。
3. 特に8世紀間に渡るスウェーデンとフィンランドの政治的な結合の間でスカンジナビアの支配を受けた時、フィンランドおよび バルト海諸国との人口交換。
●ウラル語族のY-DNA「N1c1」
Y-DNA「N1c1」はウラル語族に強く関係しており、代表的な言語系統図は下記のようになります(分類は諸説あり、それぞれ階層の 詳細はやや異なるが、属する言語に違いはない)。
サモエード語派(Nganasan語、Enets語、Nenet語およびSelkup語)。
ガナサン(Nganasan)語
エネツ(Enets)語(イェニセイ-サモエド)
ネネツ(Nenets)語
セルカップ(Selkup)語
フィン・ウゴル語派
Finno-Permic諸語
バルト・フィン諸語
フィンランド語
カレリア語
エストニア語など
ペルム(Permic)諸語
コミ(Komi)語
ウドムルト語
サーミ(Saamic)語
ヴォルガ(Volgaic)諸語
マリ(Mari)語
モルドヴィン(Mordvin)語
ウゴル諸語
ハンガリー語
Ob-ウゴル諸語
ハンティ(Khanty)語
マンシ(Mansi)語)
北シベリアのサモエード語派で最も早く分離したのは子亜型のY-DNA「N1c1*」とY-DNA「N1c1a」に一致します。
ペルム(Permic)諸語とヴォルガ(Volgaic)諸語の話し手は、Y-DNA「N1c1a1」(変異:L708)、Y-DNA「N1c1a1a」(変異:L1026)、 Y-DNA「N1c1a1a1」(変異:VL29)、Y-DNA「N1c1a1a2a」(変異:Z1935) およびY-DNA「N1c2b」(変異:P43)を含む広く様々なY-DNA「N1c」の子亜型を持っています。
バルト・フィン諸語は、Y-DNA「N1c1a1a1」(変異:VL29)がヴォルガ-ウラル地域からカレリア、フィンランドおよびエストニア への移住時に分化したように見えます。
VL29(Y-DNA「N1c1a1a1」)およびその子亜型もバルト・スラブ集団で見つかったY-DNA「N1c1」の子亜型群です、 インド・ヨーロッパ語族の亜型Y-DNA「R1a」が吸収し後に中央・東ヨーロッパのあたりでY-DNA「N1c1」から分化したことを確認します。
西シベリアのKhantyとMasiの言語と同様に、ハンガリー語を含むウゴル諸語はY-DNA「N1c1a1a2b」(変異:L1034)子亜型に相当します。
以上
表紙に戻る