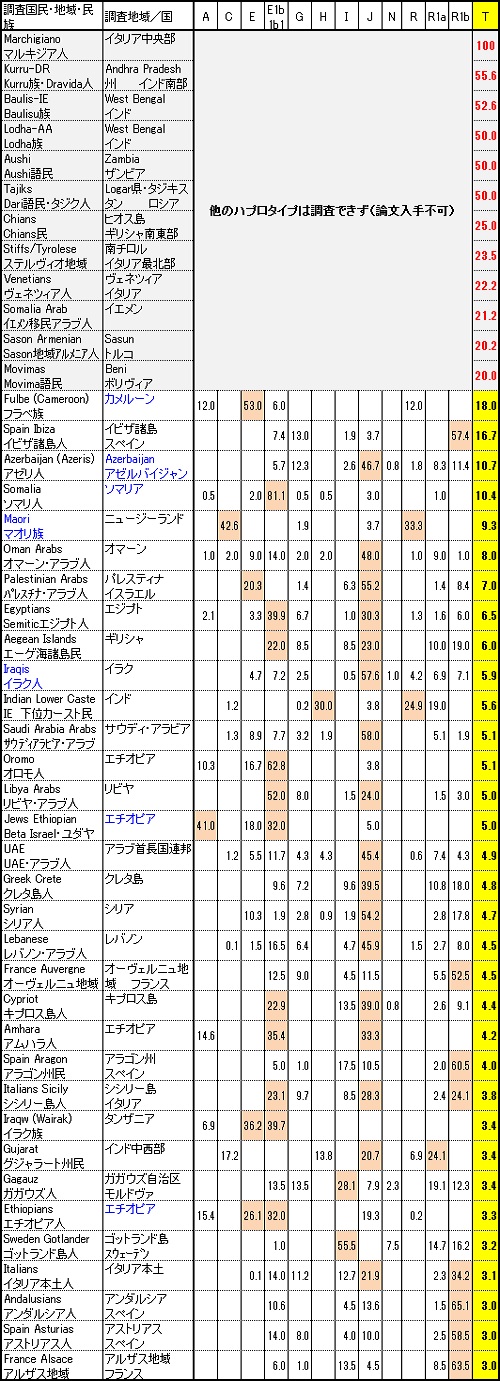
恐らくこれは第三代アメリカ大統領のトーマス・ジェファーソンが「T」と判明したため、泡沫子亜型扱いではまずいと考え独立させた ような感じを受けます。 この「T」はイタリアの中心部のMarcheという地域が「T」100%の地域であることがわかっています。民族大移動の時期に中東から他の遺伝子と 共に移動してきたのか、単独で移動してきたのかは定かではありませんが、ドラヴィダ人のKurru族で56%、インド人のBaulis族で53%を 占めるようにドラヴィダ人や交配したインド人の中に特に集中して「T」が固まって存在している独特の希少遺伝子です。 むしろ「L」の子亜型の方が自然です。
しかしこの遺伝子を含む民族を見るとヨーロッパ・中東・西アジア・北アフリカの遺伝子はほとんど構成が同じにも関わらず、 交配比率の違いと居住地の違いで外観がヨーロッパ的かセム的かに区別できます。しかしセム種が啓発した世界の3大宗教すなわち ユダヤ教、キリスト教とイスラム教の中でY-DNA「I」、「R1a」、「R1b」や「E1b1b」、「N」などのヨーロッパ人となった遺伝子が キリスト教を採用し、ほとんどのセム種やイラン種がイスラム教を採用し宗教上の違いができてしまったため、 ヨーロッパ人とセム種の違いは形質の違い以上に現在異なってしまっています。両者の元々の発祥の地域と考えられているコーカサス一帯は 両宗教が入り混じっている紛争地域でもあります。
以上
表紙に戻る
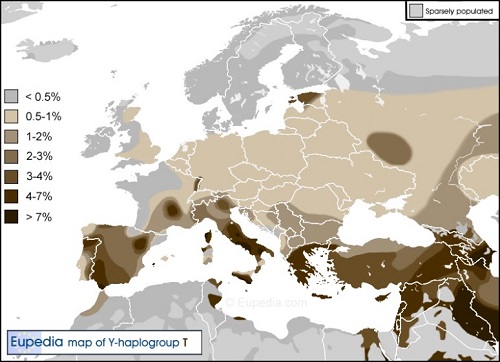
Y-DNA「T」はヨーロッパ大陸のほとんどで人口のわずか1%しか占めていません。ただし、ギリシャ、マケドニアおよびイタリアでは4%を超し、 イベリア全体では2.5%ですが、カディスでは10%、イビザでは15%以上でピークに達します。
Y-DNA「T」の最大頻度は、東アフリカ (エリトリア、エチオピア、ソマリア、ケニア、タンザニア)、および中東 (特に南コーカサス、 イラク、南西イラン、オマーンおよびエジプト南部)で、男性の血統のおよそ5?15%を占めます。
Y-DNA「T」は、これらの地方とヨーロッパに加えて、更に中央アジア、インド、カメルーン、ザンビアおよび南アフリカまで周囲から 孤立した地域でも見つかります。
その最高頻度は、実際にカメルーン (カメルーン人口の18%) のフラニ(フラベ)族で見つかります。(遊牧民が起源のフラベ族は バントゥー族、ベルベル人やアラブ人とも異なる周囲から孤立した民族)
ヨーロッパ、中東と北アフリカのY-DNA「T」の分布
Y-DNA「T」は、ユーラシアで見つけられた最も古いY-DNAの1つで少なくとも30000年前に起こりました。そのことがアフリカと 南アジア周辺の広大な分布について説明するかもしれません。
そのことはY-DNA「T」の起源地を不確かにもします。
Y-DNA「T」は、Y-DNA「K」の子亜型です (Y-DNA「K」はユーラシアのほとんどのハプロタイプ、Y-DNA「L」、「N」、「O」、「P」、 「Q」、「R」および「T」の親亜型で、起源は中東か中央アジアにあると思われています)。
Y-DNA「T」は今日、どんな場所でよりも東アフリカでより一般的ですが、農業の興隆とともにメソポタミアの肥沃な三日月地帯から 広がったことはほぼ間違いないでしょう。
確かに、Y-DNA「T」の最古の小亜型およびY-DNA「T」の最も大きな多様性は、特に中東の肥沃な三日月地帯で見つかります。
東アフリカのY-DNA「T」のより高い頻度は、中東の新石器時代の農民の間の創始者効果によるでしょう。
ヨーロッパの現代のY-DNA「T」分布は、近東の農民 (特にCardium陶磁器文化/起源前5000~1500年)による地中海ヨーロッパの 新石器時代の植民地化に強く相互に関連しています。
金石併用時代と青銅器時代の間、Y-DNA「T」は、シュメール人とエラム人のような古代の民族の間の重要な血統になっていたでしょう。
Y-DNA「T」の頻度がキプロス、シシリー、チュニジア、イビザ、アンダルシアおよびモロッコの北の先端のような場所で平均より 高いことは、フェニキア人(紀元前1200年~800年)によって地中海周辺に分散し、古代のフェニキアが、今日のレバノンにあたる地域 (フェニキア人の故郷と考えられている)よりY-DNA「T」の高い発生率を持っていたことを示唆しているようだ。
Y-DNA「T」の小亜型はほとんどすべて中東で見つかっていますが、地中海の外側のほとんどのヨーロッパ人は小亜型Y-DNA「T1a2」 (変異(L131)およびY-DNA「T1a1a」(変異P77)に属します。さらにアナトリアでも見つかります。
これらの子亜型は、恐らく肥沃な三日月地帯から南東ヨーロッパへ新石器時代の移住のうちの1つを表わしています。
その後それらは、中央および東ヨーロッパのまわりへ、東バルト海のようなはるか北へ、と広がっていったでしょう。
Y-DNA「T1a2」は、はるか東のロシアのヴォルガ-ウラル地域や北西中国の新彊で見つかりました。
Y-DNA「T」の子亜型は、恐らく新石器時代の間 (恐らくY-DNA「G2a3b1」とY-DNA「J2b2」と共に) にポントスのカスピ海の 大草原へ浸入し、青銅器時代にY-DNA「R1a」民族が中央アジアへ拡大する前に吸収されてしまったようです。
Y-DNA「T」は、ヴォルガ-ウラル地域のタタール人(5%)とマリ人(2%)の間で、同様に北西ロシア(3%)およびエストニア(3.5%)でも 比較的高頻度で見つかりました。それは、ウラル語を話す集団へ新石器時代をもたらした主要な血統のうちの1つだったかもしれない。
常染色体のDNA鑑定もまた、フィン人(1?2.5%)およびリトアニア人(1.5%)の間に、普通ではない高率の南西アジア人の混入を 同定しました。両民族はそれ以外は西アジア人かコーカサス人の混入を欠き、いかなる中東のY-DNAもほとんど持っていません。
この南西アジア人の混入は、新石器時代に吸収されたY-DNA「T」血統の名残の可能性があります。
以上
表紙に戻る