National GeographicとIBMの共同運営している「THE GENOGRAPHIC PROJECT」というサイトがありますが、氷河期の地図とホモサピエンスの拡散がわかりやすく世界地図にされているのでご参考に転載します。mtDNAの拡散図もあるのですがY-DNAの拡散図にしました。2011年以降はメンテされていないようなので最新情報ではありませんが、大まかな変化は良くわかります。
1. 6万年以前の地図です。
白い部分が氷河です。ヨーロッパと北アメリカの最北端が氷河で覆われており、ホモサピエンスの展開はまだ示されていません。
2. 55,000~60,000年前頃です。
寒冷化が始まりました。北米の半分はすっかり氷河に覆われています。
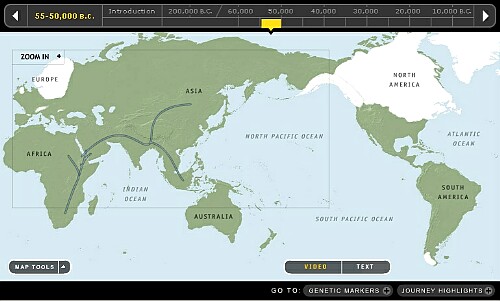
3. 50,000~55,000年前頃です。
アフリカでホモサピエンスがY-DNA「A」から「B」に分化しています。 「B」の一部が出アフリカし恐らく近東でネアンデルタール人と出会い更に「DE」と「CF」に分化し、東ユーラシア(中国大陸とスンダランド)に展開したようです。 この東ユーラシアに展開したのが古代遺伝子Y-DNA「D」と行動を共にしたとされている古代遺伝子Y-DNA「C」のようです。 Y-DNA「D」は東ユーラシアに展開した最初のホモサピエンスだったようです。
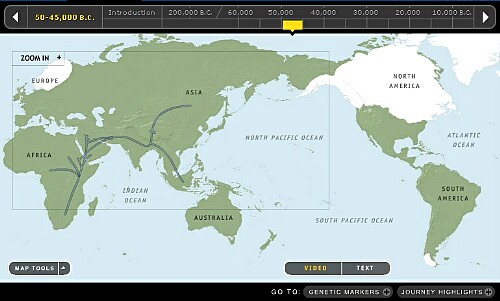
4. 45,000~50,000年前頃です。
非出アフリカ遺伝子Y-DNA「A」と「B」の子亜型への分化が進んでいます。
5. 40,000~45,000年前頃です。
インド亜大陸でY-DNA「C」は「D」との交配で子亜型に分化し、「C4」はオーストラリア大陸に既に到達していることがわかります。 同じく子亜型に分化していたY-DNA「D」はまだ中原辺りに留まっているようです。
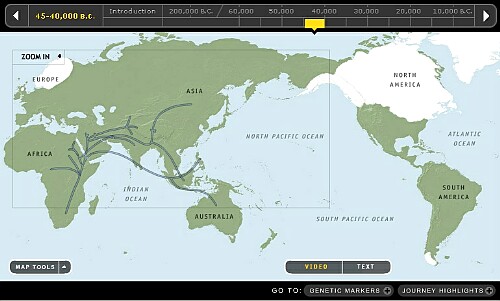
6. 35,000か~40,000年前頃です。
古代遺伝子Y-DNA「E」は環地中海域(南欧州と北アフリカ)に展開を始め、 インド亜大陸で古代遺伝子Y-DNA「F」から分化した新興遺伝子Y-DNA「I」がクロマニヨン人として欧州に移動し、兄弟遺伝子Y-DNA「J」もセム種として中近東や北アフリカに展開を始めました。 中国大陸の華南辺りではいよいよ新興遺伝子Y-DNA「O」が現れました。新興遺伝子Y-DNA「R」は更に新しい遺伝子なのでまだ現れてはいないようです。
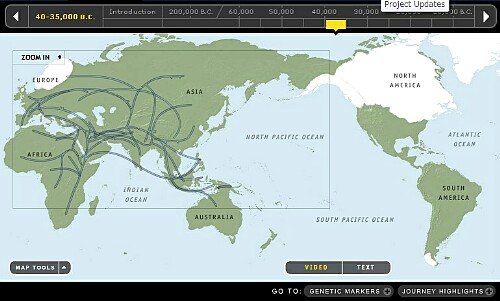
7. 30,000~35,000年前頃です。
Y-DNA「D2」が日本列島に到達したことを示しています。日本列島の旧石器時代はY-DNA「D2」及び行動を共にしていたとされているY-DNA「C1」によって確立されたと欧米では考えています。
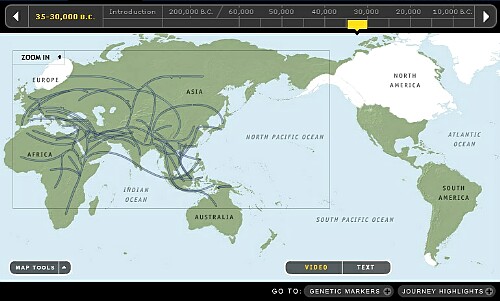
8. 25,000~30,000年前頃です。
最寒冷期(LGM)になり、北欧の氷河が大きく発達し海面がかなり降下しベーリング海峡は広大なベーリング大陸となり、Y-DNA「Q」がまだ氷河におおわれている北米大陸に到達しました。 この時に4タイプ程度のmtDNAも同時に渡っています。日本列島にはY-DNA「O3」が到達したようになっていますが勿論間違いです。この頃に渡ってきたとするならY-DN「C3a」のはずです。
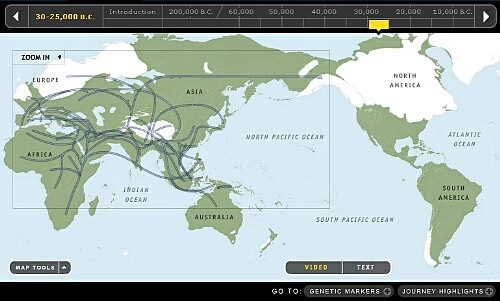
9. 20,000~25,000年前頃です。
Y-DNA「C3a」は最寒冷化するシベリアから逃げ南下してきた大型獣を追って日本列島に入ったようです。 一方北東沿岸沿いに移動した部隊Y-DNA「C3b」はとうとうベーリング大陸を渡り北米に到達しY-DNA「Q」と共にネイティヴアメリカンの一部になりました。 中米で発見される縄文土器似の土器の製作者のようです。
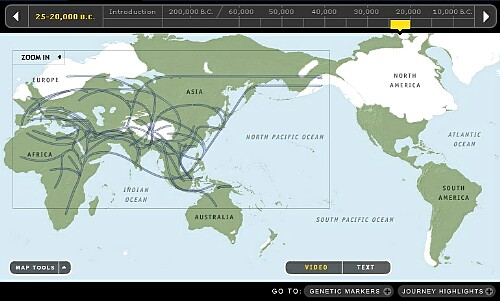
10. 15,000~20,000年前頃です。
Y-DNA「Q」はとうとう南米大陸に到達しました。しかしY-DNA「C3b」は中米に留まり南米大陸には南下しなかったようです。
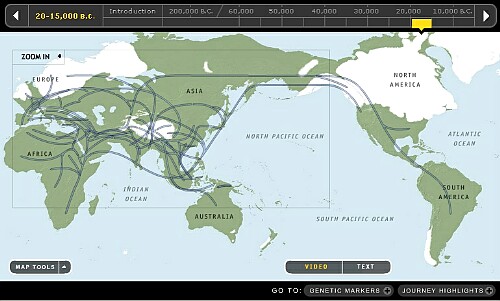
11. 10,000~15,000年前頃です。
氷河の縮退が始まりました。北欧を覆っていた氷河はスカンジナビアのみに縮小し、北ヒマラヤの氷河も消失しました。
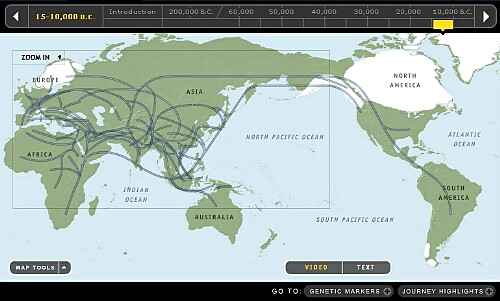
12. ~10,000年前頃です。
間氷期になり温暖化し北極圏以外の氷河は消失し、現在に続いています。海面は上昇し現在の海岸線が出来上がり、更なる温暖化で海面は徐々に上昇し続けています。
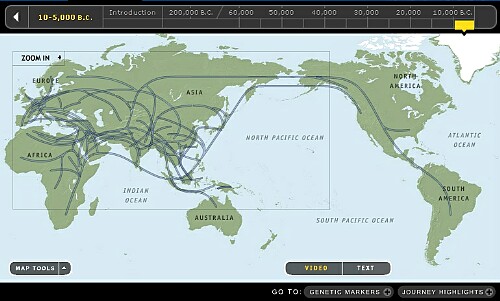
ご参考にmtDNAの軌跡図です。
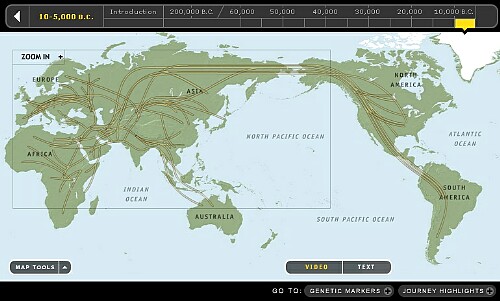
古代の女性は集団を越えて他の集団に移動するという、人類とチンパンジーにのみ認められる行動原理で先ず女性に現れる新しい遺伝子を拡散してきました。
出アフリカせずにアフリカに残留していたホモエレクトスからネアンデルタール人が進化し出アフリカをはたしましたが、 その間ホモエレクトスのまま進化できなかった残留組からやっとmtDNAイヴが最新の学説で20万年前頃(前説では14万年前頃)に誕生して、 6万年も後の14万年前頃(前説では9万年前頃)にやっとY-DNAアダムが誕生したくらいホモサピエンスのY-DNAが確立するには時間がかかっているのです。 最新学説では14万年前頃にY-DNAアダムが誕生してハプロタイプY-DNA「A」になるわけですが、次のY-DNA「B」が分化するのは最新説も前説と変わらず60000~65000年前頃ごろだそうです。
とにかく進化はかなり時間がかかっているのです(と言っても地球年齢から見るとほんの秒単位にもならないそうですが...)。 つまり人類が進化を始めるのに8万年近く(前説では3万年近く)かかっているのです。人類はホモサピエンスに進化してもすぐに古代遺伝子「C」「D」「E」「F」が分化してきたわけではないのです。 「B」の一部が乾燥化して住めなくなったサハラ砂漠から絶滅危惧種に陥りながら先輩人類同様出アフリカを果たし、近東で先輩の旧人ネアンデルタール人と遭遇し、 交配し初めて古代遺伝子「DE」と「CF」に分化を果たしたのです。
もしネアンデルタール人と遭遇・交配をしていなかったら50000年前頃一気に高まった現代人類の祖先の技術力や芸術性などの進歩はもっと遅れて、 農業革命も遅れ人類は出アフリカしなかったコイサン族やピグミー族などと同様に狩猟採集の今だ古代のままだった可能性も充分にあるのです。 ところが、インド亜大陸で恐らく地の先住ネアンデルタール人と更に交配が進み「CF」から古代遺伝子「C」と「F」が分化し、「F」が更に「G」以降の新興遺伝子群に分化することが出来たおかげで、 ホモサピエンスは更に進化する可能性を手に入れたようです。
この時に「DE」も「D」と「E」に分化したのですが、「D」はアンダマン諸島のOnge族のように狩猟採集のまま留まり、「E」はこともあろうにアフリカに戻ってしまったわけです。 結局残念ながら「D」も「E」も「CF」から分化した一方の「C」も新興遺伝子に分化することはできませんでした。
まとめると日本人や日本文化の基層にあるのはY-DNA「D2」「C1a」「C3a」が1万年に渡って日本列島で展開してきた狩猟採集(晩期には農耕もあったようだ)の縄文文化であり、 チベット古語と共通する日本語の基層の文法を構成する縄文語です。そして1万年後にやってきた呉系長江文化弥生人Y-DNA「O2b」「O2a」「O1a」と縄文遺伝子が交配し 分化した弥生主役の「O2b1」が新たな農耕文化を日本列島に定着させ、縄文系と密に交配し日本列島を瑞穂の国にしたのです。
そして最後に朝鮮半島から生き残り競争に負けて追い出されてきた武装侵攻集団「O3」が日本列島を支配し大和朝廷や武士団内で内部抗争を繰り返し、 明治維新を経て戦後ようやくエスタブリッシュメントの「O3」と他の先住遺伝子の交配は進みつつありますが、まだまだ日本列島独自の「O3」子亜型が生じるまでには至っていません。 「O3」は今でも政治や経済の支配階級でもあるのです。田舎に多い縄文-弥生の交配集団は黙々と従う側なのです。
以上
表紙に戻る
